Гладкая мышечная ткань образует мышечную оболочку трубкообразных органов пищеварения, дыхания, выделения, размножения, находится в стенках кровеносныхсосудов, протоков желез, в селезенке, коже и других органах.
Специализированные сократительные гладкомышечные ткани входят в состав потовых, слюнных, молочных желез. Сократительные клетки этих желез в своей цитоплазме содержат миофиламенты, построенные из сократительных белков и развиваются из эпителиальных клеток. Другие разновидности специализированных сократительных тканей имеют нейроглиальное происхождение, суживают зрачок и располагаются в радужной оболочке глаза.
Гладкая мышечная ткань относится к ткани с непроизвольным сокращением, её функцию контролирует вегетативная нервная система. Сокращения гладких мышц могут быть медленными, но достигать большой силы сжатия.
Основной структурной единицей гладкой мышечной ткани являются клетки-миоциты. Они удлиненной веретеновидной формы с заостренными концами. Их длина от 20 до 200 мкм (в беременной матке до 500 мкм), а толщина 8-10 мкм. Ядро палочковидной формы находится в середине клетки. В цитоплазме, около полюсов ядра расположены органеллы: митохондрии, комплекс Гольджи, центросома, рибосомы, эндоплазматическая сеть и включения гликогена (энергетический резерв клетки). В преферической части цитоплазмы расположены миофиламенты. Нити актина и миозина не образуют миофибрилл или постоянных акто-миозиновых комплексов и расположены по-разному. Актиновые нити чаще имеют косое присоединение к плазмолемме с помощью особых плотных телец (прикрепительных дисков). Отдельные пучки актиновых нитей прикрепляются к плотным тельцам, расположенным в цитоплазме. Положение этих телец или дисков с обратной стороны закрепляется промежуточными филаментами.
Миозиновые нити в периоды расслабления миоцитов лежат в цитоплазме продольно или под углом к длинной оси клетки. В процессе сокращения актиновые и миозиновые нити смещаются навстречу друг другу и формируют акто-миозиновые комплексы. В результате клетка сокращается и приобретает неправильную форму. В фазе расслабления комплексы вновь распадаются. Поскольку актиновые и миозиновые нити лежат неупорядоченно, поперечная исчерченность в гладких миоцитах отсутствует.
В процессе сокращения, как было сказано, важную роль играют ионы Са ++ . Депо для них является гладкая эндоплазматическая сеть миоцита. Кроме того, ионы Са ++ поступают извне через кальциевые каналы в цитолемме. В определенных участках плазмолеммы лежат специальные белки, воспринимающие и пропускающие внутрь ионы Са ++ . Ионы Са ++ в комплексе с белком кольмодулином и ферментом киназой запускают процесс сокращения. Головки молекул миозина начинают двигаться и скользить вдоль нитей актина и осуществляется процесс сокращения.
С помощью электронного микроскопа было выявлено, что на концах гладких миоцитов имеются пальцевидные выпячивания, десмосомы и щелевидные контакты-нексусы. Плазмолемма миоцитов впячиваясь в цитоплазму, образует пузырьки (кавеолы), примыкающие к саркоплазматической сети. Предполагают, что эти пузырьки участвуют в проведении нервных импульсов, вызывающих выход ионов Са ++ и процесс сокращения.
Функциональной единицей гладкой мышечной ткани является пучок из 10-15 миоцитов, связанных с одним нервным волокном. Благодаря тесной связи клеток с помощью десмосом и щелевых контактов все клетки пучка быстро реагируют на нервное раздражение, несмотря на то, что нервное окончание входит только в одну клетку.
Коллагеновые волокна, соединительно-тканных капсул (эндомизий) оплетают миоциты, вплетаются в базальную пластинку (мембрану) в наружный слой над сарколеммой и тем самым удерживают клетки от чрезмерного сжатия и растяжения.
Пучки отделены друг от друга прослойками соединительной ткани (перемизий), в которой проходят сосуды и нервы.
Гладкая мышечная ткань, иннервируется вегетативной нервной системой. Ее деятельность регулируется корой полушарий, но без участия сознания. Сокращения осуществляются непроизвольно и происходят медленно и ритмично (период сокращения от 3 до 5 минут).
Такой характер сокращения называют тоническим .
В стенках полых органов и сосудов пучки гладких миоцитов объединяются в пласты (продольные и циркулярные) .
Гладкая мышечная ткань обладает большой силой, передвигая в кишечнике большие массы пищи, и обладает слабой утомляемостью. В стенке кишечника сокращение происходит 12 раз в минуту.
Отдельные пучки гладких миоцитов находятся в коже животных в виде мышц, поднимающих волос.
Происходит гладкая мышечная ткань внутренних органов и сосудов из мезенхимы, клетки которой дифференцируются в миобласты, а миобласты в миоциты, сохраняющие способность к делению на протяжении всего онтогенеза. Кроме того, гладкие миоциты могут образовываться из недеффиренцированных клеток соединительной ткани (адвентициальных), находящихся около кровеносных сосудов.
Мышечная ткань (textus muscularis) представляет собой группу тканей (поперечнополосатая, гладкая, сердечная), имеющих различное происхождение и строение, объединенных по функциональному признаку - способности сокращаться - укорачиваться. Наряду с упомянутыми разновидностями мышечной ткани, образующейся из мезодермы (мезенхимы), в организме человека выделяют мышечную ткань эктодермального происхождения - миоциты радужки глаза.
Исчерченная (поперечнополосатая, скелетная) мышечная ткань образована цилиндрическими мышечными волокнами длиной от 1 мм до 4 см и более и толщиной до 0,1 мм. Каждое волокно представляет собой комплекс, состоящий из миосимпласта и миосателлитоцитов, покрытых плазматической мембраной, которую называют сарколеммой (от греч. sarkos - мясо). Снаружи к сарколемме прилежит базальная пластинка (мембрана), образованная тонкими коллагеновыми и ретикулярными волокнами. Миосимпласт, находящийся под сарколеммой мышечного волокна, получил название саркоплазмы. Он состоит из множества эллипсоидных ядер (до 100), миофибрилл и цитоплазмы. Удлиненные ядра, ориентированные вдоль мышечного волокна, лежат под сарколеммой. В саркоплазме имеется большое количество элементов зернистой эндоплазматической сети. Примерно 1/3 сухой массы мышечного волокна приходится на цилиндрические миофибриллы, проходящие продольно почти через всю саркоплазму. Между миофибриллами располагаются многочисленные митохондрии с хорошо развитыми кристами и гликоген.
В поперечнополосатом мышечном волокне хорошо развита саркотубулярная сеть, которая образована двумя компонентами: канальцами эндоплазматической сети, расположенными вдоль миофибрилл (L-система), и Т-канальцами (Т-трубочками), начинающимися в области впячивания сарколеммы. Т-трубочки проникают в глубь мышечного волокна и образуют поперечные трубочки вокруг каждой миофибриллы.
Т-трубочки играют важную роль в быстром проведении потенциала действия к каждой миофибрилле. Возникший в сарколемме мышечного волокна под воздействием нервного импульса потенциал действия распространяется по Т-трубочкам, а от них на незернистую эндоплазматическую сеть, канальцы которой располагаются возле Т-трубочек, а также между миофибриллами.
Основную часть саркоплазмы мышечного волокна составляют специальные органеллы - миофибриллы. Каждая миофибрилла состоит из правильно чередующихся участков - темных анизотропных дисков А и светлых изотропных дисков I. В середине каждого анизотропного диска А проходит светлая зона - полоска Н, в центре которой находится линия М, или мезофрагма. Через середину диска I проходит линия Z - так называемая телофрагма. Чередование темных и светлых дисков в соседних миофибриллах, располагающихся на одном уровне, на гистологическом препарате скелетной мышцы создает впечатление поперечной исчерченности. Каждый темный диск образован толстыми миозиновыми нитями диаметром 10-15 нм. Длина толстых нитей около 1,5"мкм. Основу этих нитей (филаментов) составляет высокомолекулярный белок миозин. Каждый светлый диск образован из тонких актиновых нитей диаметром 5-8 нм и длиной около 1 мкм, состоящих из низкомолекулярного белка актина, а также низкомолекулярных белков тропомиозина и тропонина.
Участок миофибриллы между двумя телофрагмами (Z-линиями) называют саркомером. Он является функциональной единицей миофибриллы. Длина саркомера около 2,5 мкм, в него входят темный диск А и половинки примыкающих к нему с двух сторон светлых дисков I. Таким образом, тонкие актиновые нити идут от Z-линии навстречу друг другу и входят в диск А, в промежутки между толстыми миозиновыми нитями. При сокращении мышцы актиновые и миозиновые нити скользят навстречу друг другу, при расслаблении - двигаются в противоположные стороны.
Саркоплазма богата белком миоглобином, который, подобно гемоглобину, может связывать кислород. В зависимости от толщины волокон, содержания в них миоглобина и миофибрилл различают так называемые красные и белые поперечнополосатые мышечные волокна. Красные мышечные волокна (темные) богаты саркоплазмой, миоглобином и митохондриями, однако в них мало миофибрилл. Эти волокна медленно сокращаются и долго могут быть в сокращенном (рабочем) состоянии. Белые мышечные (светлые) волокна содержат мало саркоплазмы, миоглобина и митохондрий, но в них много миофибрилл. Эти волокна сокращаются быстрее красных, но быстро «устают». У человека мышцы содержат оба типа волокон. Сочетание медленных (красных) и быстрых (белых) мышечных волокон обеспечивает мышцам быстроту реакции (сокращение) и длительную работоспособность.
Миосателлитоцит ы располагаются непосредственно над сарколеммой, но под базальной пластинкой (мембраной). Это уплощенные клетки с богатым хроматином крупным ядром. Каждый миосателлитоцит имеет центросому и малое число органелл; спиральных органелл (миофибрилл) у них нет. Миосателлитоциты являются стволовыми (ростковыми) клетками поперечнополосатой (скелетной) мышечной ткани, они способны к синтезу ДНК и митотическому делению.
Неисчерчеиная (гладкая) мышечная ткань состоит из миоцитов, которые располагаются в стенках кровеносных, лимфатических сосудов, полых внутренних органов, где образуют их сократительный аппарат. Гладкие миоциты - это удлиненные веретенообразные клетки длиной от 20 до 500 мкм и толщиной от 5 до 15 мкм, лишенные поперечной исчерченности. Клетки располагаются группами, заостренный конец каждой клетки внедряется между двумя соседними клетками. Каждый миоцит окружен базальной мембраной, коллагеновыми и ретикулярными волокнами, среди которых проходят эластические волокна. Клетки связаны между собой многочисленными нексусами. Удлиненное палочковидное ядро, достигающее 10-25 мкм в длину, при сокращении клетки принимает форму штопора. Изнутри к цитолемме прилежат расположенные в цитоплазме веретенообразные плотные (прикрепительные) тельца.
Плотные тельца являются аналогом Z-полосок поперечнополосатых мышечных волокон. В их состав входит белок а-актинин.
В цитоплазме гладких миоцитов находятся миофиламенты двух типов - тонкие и толстые. Тонкие актиновые миофиламенты диаметром 3-8 нм лежат вдоль миоцита или косо по отношению к его длинной оси. Они прикрепляются к плотным тельцам. Толстые короткие миозиновые миофиламенты диаметром около 15 нм располагаются в цитоплазме продольно. Тонкие и толстые нити не образуют саркомеров, поэтому гладкие миоциты не имеют поперечной исчерченности. При сокращении миоцитов актиновые и миозиновые миофиламенты смещаются навстречу друг другу, при этом гладкомышечная клетка укорачивается.
Группа миоцитов, окруженных соединительной тканью, иннервируется обычно одним нервным волокном. Нервный импульс передается с одной мышечной клетки на другую благодаря нексусам со скоростью 8-10 см/с. В некоторых гладких мышцах (например, сфинктер зрачка) иннервируется каждый миоцит.
Скорость сокращения гладких миоцитов значительно меньше, чем поперечнополосатых мышечных волокон (в 100-1000 раз), при этом гладкими миоцитами затрачивается в 100-500 раз меньше энергии.
Гладкие мышцы совершают длительные тонические сокращения (например, сфинктеры полых - трубчатых - органов, гладкие мышцы кровеносных сосудов) и относительно медленные движения, которые часто ритмичные.
Исчерченная сердечная мышечная ткань относится к поперечнополосатой, однако по своему строению и функции отличается от скелетных мышц. Она состоит из сердечных миоцитов (кардиомиоцитов), образующих соединяющиеся друг с другом комплексы. Сокращения сердечной мышцы не контролируются сознанием человека. Кардиомиоциты - это клетки неправильной цилиндрической формы длиной 100-150 мкм и диаметром 10-20 мкм. Каждый кардиомиоцит имеет один-два овальных ядра, лежащих в центре и окруженных миофибриллами, расположенными по периферии строго продольно. Возле обоих полюсов ядра видны зоны цитоплазмы, лишенные миофибрилл. Строение миофибрилл в кардиомиоцитах аналогично их строению в скелетных мышцах. В кардиомиоцитах большое количество крупных митохондрий с хорошо развитыми кристами, которые располагаются группами между миофибриллами. Под цитолеммой и между миофибриллами находятся гликоген и структуры незернистой эндоплазматической сети. Эта сеть образует канальцы L-системы, с которыми контактируют Т-трубочки.
Кардиомиоциты соединены между собой так называемыми вставочными дисками, которые при светооптическом изучении имеют вид темных полосок. Вставочный диск - это зона контакта двух кардиомиоцитов, включающая цитолемму этих клеток, десмосомы, нексусы и зоны прикрепления миофибрилл каждого кардиомиоцита к его цитолемме. Десмосомы и нексусы соединяют соседние кардиомиоциты друг с другом. Через нексусы происходят передача нервного возбуждения и обмен ионами между клетками.
Различают три группы гладких (неисчерченных) мышечных тканей (textus muscularis nonstriatus) и клеток: мезенхимные, нейральные и миоэпи-телиальные клетки.
9.3.1. Мышечная ткань мезенхимного происхождения
Гистогенез. Эта ткань делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоци-тов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их в цитоплазме становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние. Дифференцируясь, они синтезируют компоненты межклеточного матрикса, коллаген базальной мембраны, а также эластин. У дефинитивных клеток (миоцитов) синтетическая способность снижена, но не исчезает полностью.
Строение и функционирование клеток. Гладкий миоцит - веретеновидная клетка длиной 20-500 мкм, шириной 5-8 мкм. Ядро палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается (рис. 9.12-9.14).
Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерии обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм. Наибольшей длины гладкие мио-
циты достигают в стенке матки - до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактильные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность.
Секреторные миоциты по своей ультраструктуре напоминают фиб-робласты, однако содержат в своей цитоплазме пучки тонких миофи-ламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцирован-ным. Филаменты актина образуют в цитоплазме трехмерную сеть, вытянутую преимущественно продольно. Концы филаментов скреплены между собой и с плазмолеммой специальными сшивающими белками. Эти участки хорошо видны на электронных микрофотографиях как плотные тельца. Мономеры миозина располагаются рядом с филаментами актина. Плазмолемма образует впя-чивания - кавеолы, в которых концентрируются ионы кальция. Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их термина-лей, изменяет проницаемость плаз-молеммы. Происходит высвобождение ионов кальция, что влечет за собой как полимеризацию миозина, так и взаимодействие миозина с актином.
Происходит втягивание актино-вых миофиламентов между миози-
Рис. 9.12. Строение гладкого миоцита (схема):
а, в - при расслаблении; б, д - при наибольшем сокращении; г - при неполном сокращении; в-д - увеличенные изображения участков, обведенных рамками на фрагментах а и б. 1 - плазмолемма; 2 - плотные тельца; 3 - ядро; 4 - эндоплазма; 5 - сократительные комплексы; 6 - митохондрии; 7 - базальная мембрана; 8 - актиновые (тонкие) мио-филаменты; 9 - миозиновые (толстые) миофиламенты
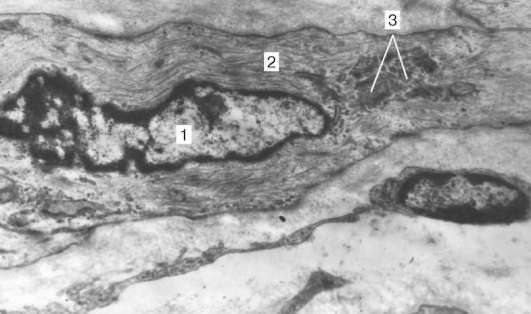
Рис. 9.13. Ультраструктура дифференцирующегося гладкого миоцита в стенке бронха:
1 - ядро; 2 - цитоплазма с миофиламентами; 3 - комплекс Гольджи, увеличение 35 000 (препарат А. Л. Зашихина)
новыми, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается (см. рис. 9.12). Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция перемещаются из цитоплазмы в кавеолы и в канальцы эндоплазматической сети, миозин деполимеризуется и «миофибриллы» распадаются. Сокращение прекращается. Таким образом, актиномиозиновые комплексы существуют в гладких миоцитах только в период сокращения при наличии в цитоплазме свободных ионов кальция.
Миоциты окружены базальной мембраной. На отдельных участках в ней образуются «окна», поэтому плазмолеммы соседних миоцитов сближаются. Здесь формируются нексусы, и между клетками возникают не только механические, но и метаболические связи. Поверх «чехликов» из базальной мембраны между миоцитами проходят эластические и ретикулярные волокна, объединяющие клетки в единый тканевой комплекс. Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и аморфный компонент межклеточного матрикса. Взаимодействие миоцитов осуществляется при помощи цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом, участков мембранных контактов поверхностей миоцитов.
Регенерация. Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов. В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазматического отношения, увеличение количества свободных рибосом и полисом, актива-

Рис. 9.14. Строение гладкой мышечной ткани (объемная схема) (по Р. В. Крстичу, с изменениями):
1 - веретеновидные гладкие миоциты; 2 - цитоплазма миоцита; 3 - ядра миоци-тов; 4 - плазмолемма; 5 - базальная мембрана; 6 - поверхностные пиноцитозные пузырьки; 7 - межклеточные соединения; 8 - нервное окончание; 9 - коллагеновые фибриллы; 10 - микрофиламенты
ция ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема). При действии ряда повреждающих факторов отмечается феноти-пическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании интимальной гиперплазии при развитии атеросклероза.
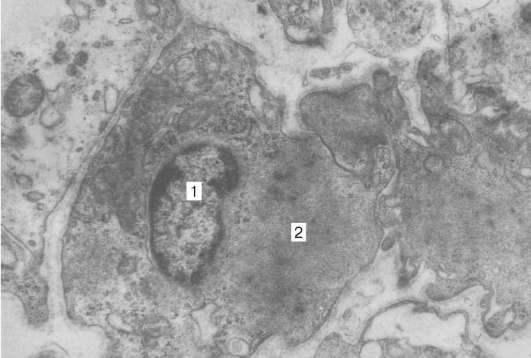
Рис. 9.15. Ультраструктура миопигментоцита (препарат Н. Н. Сарбаевой): 1 - ядро; 2 - миофиламенты, увеличение 6000
9.3.2. Мышечная ткань мезенхимного типа в составе органов
Миоциты объединяются в пучки, между которыми располагаются тонкие прослойки соединительной ткани. В эти прослойки вплетаются ретикулярные и эластические волокна, окружающие миоциты. В прослойках проходят кровеносные сосуды и нервные волокна. Терминали последних оканчиваются не непосредственно на миоцитах, а между ними. Поэтому после поступления нервного импульса медиатор распространяется диффуз-но, возбуждая сразу многие клетки. Гладкая мышечная ткань мезенхимно-го происхождения представлена главным образом в стенках кровеносных сосудов и многих полых внутренних органов.
Гладкая мышечная ткань в составе конкретных органов имеет неодинаковые функциональные свойства. Это обусловлено тем, что на поверхности органов имеются разные рецепторы конкретных биологически активных веществ. Поэтому и на многие лекарственные препараты их реакция неодинакова. Возможно, разные функциональные свойства тканей связаны и с конкретной молекулярной организацией актиновых филаментов.
9.3.3. Мышечная ткань нейрального происхождения
Мышечная ткань радужки и цилиарного тела относится к четвертому типу сократимых тканей. Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. В ряду
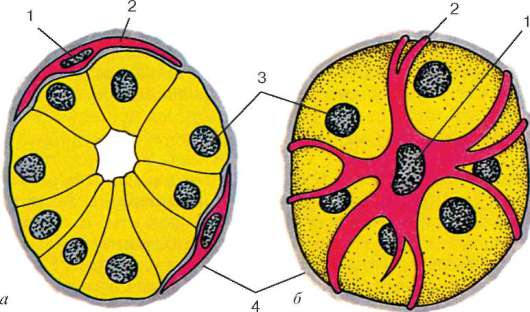
Рис. 9.16. Миоэпителиальные клетки в концевом отделе слюнной железы (схема по Г. С. Катинасу):
а - поперечный срез; б - вид с поверхности. 1 - ядра миоэпителиоцитов; 2 - отростки миоэпителиоцитов; 3 - ядра секреторных эпителиоцитов; 4 - базальная мембрана
позвоночных мышечные элементы радужки обнаруживают разнообразную дивергентную дифференцировку. Так, мионейральная ткань у рептилий и птиц представлена исчерченными многоядерными волокнами, имеющими большое сходство с мускулатурой скелетного типа. У млекопитающих и человека основной структурно-функциональной единицей мышц радужки является гладкий одноядерный миоцит, или миопигментоцит. Последние имеют пигментированное тело, содержащее одно ядро, вынесенное за пределы веретеновидной сократимой части (рис. 9.15).
Цитоплазма клеток содержит большое число митохондрий и пигментные гранулы, которые сходны по размерам и форме с гранулами пигментного эпителия. Миофиламенты в миопигментоцитах делятся на тонкие (7 нм) и толстые (1,5 нм), по размерам и расположению напоминают миофиламен-ты гладких миоцитов. Каждый миопигментоцит окружен базальной мембраной. Возле цитоплазматических отростков миоцитов обнаруживаются безмиелиновые нервные волокна. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы - суживающую и расширяющую зрачок.
Регенерация. В немногочисленных работах показана низкая регенераци-онная активность после повреждения или ее отсутствие.
9.3.4. Мышечные клетки эпидермального происхождения
Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с их секреторными клетками. Миоэпителиальные
клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки восстанавливаются тоже из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез (рис. 9.16). В теле клетки располагаются ядро и органеллы общего значения, а в отростках - сократительный аппарат, организованный как и в клетках мышечной ткани мезенхимного типа.
Контрольные вопросы
1. Генетическая классификация мышечных тканей. Структурно-функциональные единицы разных типов мышечной ткани.
2. Поперечнополосатая скелетная мышечная ткань: развитие, строение, морфологические основы мышечного сокращения. Регенерация.
3. Поперечнополосатая сердечная мышечная ткань: развитие, специфика строения различных видов кардиомиоцитов, регенерация.
4. Разновидности гладких миоцитов: источники развития, топография в организме, регенерация.







