Подвижность является характерным свойством всех форм жизни. Направленное движение имеет место при расхождении хромосом в процессе клеточного деления, активном транспорте молекул, перемещении рибосом в ходе белкового синтеза, сокращении и расслаблении мышц. Мышечное сокращение – наиболее совершенная форма биологической подвижности. В основе любого движения, в том числе и мышечного, лежат общие молекулярные механизмы.
У человека различают несколько видов мышечной ткани. Поперечно-полосатая мышечная ткань составляет мышцы скелета (скелетные мышцы, которые мы можем сокращать произвольно). Гладкая мышечная ткань входит в состав мышц внутренних органов: желудочно-кишечного тракта, бронхов, мочевыводящих путей, кровеносных сосудов. Эти мышцы сокращаются непроизвольно, независимо от нашего сознания.
В данной лекции мы рассмотрим строение и процессы сокращения и расслабления скелетных мышц, поскольку именно они представляют наибольший интерес для биохимии спорта.
Механизм мышечного сокращения до настоящего времени раскрыт не полностью.
Достоверно известно следующее.
1. Источником энергии для мышечного сокращения являются молекулы АТФ.
2. Гидролиз АТФ катализируется при мышечном сокращении миозином, обладающим ферментативной активностью.
3. Пусковым механизмом мышечного сокращения является повышение концентрации ионов кальция в саркоплазме миоцитов, вызываемое нервным двигательным импульсом.
4. Во время мышечного сокращения между тонкими и толстыми нитями миофибрилл возникают поперечные мостики или спайки.
5. Во время мышечного сокращения происходит скольжение тонких нитей вдоль толстых, что приводит к укорочению миофибрилл и всего мышечного волокна в целом.
Гипотез объясняющих механизм мышечного сокращения много, но наиболее обоснованной является так называемая гипотеза (теория) «скользящих нитей» или «гребная гипотеза».
В покоящейся мышце тонкие и толстые нити находятся в разъединенном состоянии.
Под воздействием нервного импульса ионы кальция выходят из цистерн саркоплазматической сети и присоединяются к белку тонких нитей – тропонину. Этот белок меняет свою конфигурацию и меняет конфигурацию актина. В результате образуется поперечный мостик между актином тонких нитей и миозином толстых нитей. При этом повышается АТФазная активность миозина. Миозин расщепляет АТФ и за счет выделившейся при этом энергии миозиновая головка подобно шарниру или веслу лодки поворачивается, что приводит к скольжению мышечных нитей навстречу друг другу.
Совершив поворот, мостики между нитями разрываются. АТФазная активность миозина резко снижается, прекращается гидролиз АТФ. Однако при дальнейшем поступлении нервного импульса поперечные мостики вновь образуются, так как процесс, описанный выше, повторяется вновь.
В каждом цикле сокращения расходуется 1 молекула АТФ.
В основе мышечного сокращения лежат два процесса:
спиральное скручивание сократительных белков;
циклически повторяющееся образование и диссоциация комплекса между цепью миозина и актином.
Мышечное сокращение инициируется приходом потенциала действия на концевую пластинку двигательного нерва, где выделяется нейрогормон ацетилхолин, функцией которого является передача импульсов. Сначала ацетилхолин взаимодействует с ацетилхолиновыми рецепторами, что приводит к распространению потенциала действия вдоль сарколеммы. Все это вызывает увеличение проницаемости сарколеммы для катионов Na + , которые устремляются внутрь мышечного волокна, нейтрализуя отрицательный заряд на внутренней поверхности сарколеммы. С сарколеммой связаны поперечные трубочки саркоплазматического ретикулума, по которым распространяется волна возбуждения. От трубочек волна возбуждения передается мембранам пузырьков и цистерн, которые оплетают миофибриллы на участках, где происходит взаимодействие актиновых и миозиновых нитей. При передаче сигнала на цистерны саркоплазматического ретикулума, последние начинают освобождать находящийся в них Са 2+ . Высвобожденный Са 2+ связывается с Тн-С, что вызывает конформационные сдвиги, передающиеся на тропомиозин и далее на актин. Актин как бы освобождается из комплекса с компонентами тонких филаментов, в котором он находился. Далее актин взаимодействует с миозином, и результатом такого взаимодействия является образование спайки, что делает возможным движение тонких нитей вдоль толстых.
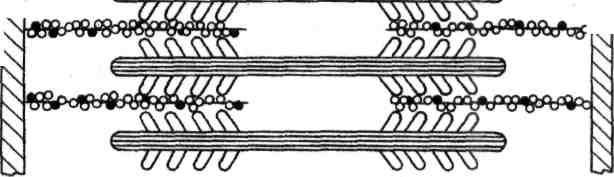
Генерация силы (укорочение) обусловлена характером взаимодействия между миозином и актином. На миозиновом стержне имеется подвижный шарнир, в области которого происходит поворот при связывании глобулярной головки миозина с определенным участком актина. Именно такие повороты, происходящие одновременно в многочисленных участках взаимодействия миозина и актина, являются причиной втягивания актиновых филаментов (тонких нитей) в Н-зону. Здесь они контактируют (при максимальном укорочении) или даже перекрываются друг с другом, как это показано на рисунке.

 в
в
Рисунок. Механизм сокращения: а – состояние покоя; б – умеренное сокращение; в – максимальное сокращение
Энергию для этого процесса поставляет гидролиз АТФ. Когда АТФ присоединяется к головке молекулы миозина, где локализован активный центр миозиновой АТФазы, связи между тонкой и толстой нитями не образуется. Появившийся катион кальция нейтрализует отрицательный заряд АТФ, способствуя сближению с активным центром миозиновой АТФазы. В результате происходит фосфорилирование миозина, т. е. миозин заряжается энергией, которая используется для образования спайки с актином и для продвижения тонкой нити. После того как тонкая нить продвинется на один «шаг», АДФ и фосфорная кислота отщепляются от актомиозинового комплекса. Затем к миозиновой головке присоединяется новая молекула АТФ, и весь процесс повторяется со следующей головкой молекулы миозина.
Затрата АТФ необходима и для расслабления мышц. После прекращения действия двигательного импульса Са 2+ переходит в цистерны саркоплазматического ретикулума. Тн-С теряет связанный с ним кальций, следствием этого являются конформаци-онные сдвиги в комплексе тропонин-тропомиозин, и Тн-I снова закрывает активные центры актина, делая их неспособными взаимодействовать с миозином. Концентрация Са 2+ в области сократительных белков становится ниже пороговой, и мышечные волокна теряют способность образовывать актомиозин.
В этих условиях эластические силы стромы, деформированной в момент сокращения, берут верх, и мышца расслабляется. При этом тонкие нити извлекаются из пространства между толстыми нитями диска А, зона Н и диск I приобретают первоначальную длину, линии Z отдаляются друг от друга на прежнее расстояние. Мышца становится тоньше и длиннее.
Скорость гидролиза АТФ при мышечной работе огромна: до 10 мк моль на 1 г мышцы за 1 мин. Общие запасы АТФ невелики, поэтому для обеспечения нормальной работы мышц АТФ должна восстанавливаться с той же скоростью, с какой она расходуется.
Расслабление мышцы происходит после прекращения поступления длительного нервного импульса. При этом проницаемость стенки цистерн саркоплазматической сети уменьшается, и ионы кальция под действием кальциевого насоса, используя энергию АТФ, уходят в цистерны. Удаление ионов кальция в цистерны ретикулума после прекращения двигательного импульса требует значительных энерготрат. Так как удаление ионов кальция происходит в сторону более высокой концетрации, т.е. против осмотического градиента, то на удаление каждого иона кальция затрачивается две молекулы АТФ. Концентрация ионов кальция в саркоплазме быстро снижается до исходного уровня. Белки вновь приобретают конформацию характерную для состояния покоя.
Таким образом, и процесс мышечного сокращения и процесс мышечного расслабления – это активные процессы, идущие с затратами энергии в виде молекул АТФ,
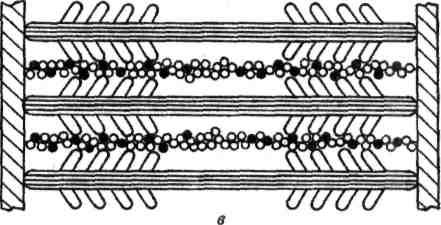
В гладких мышцах нет миофибрилл, которые состоят из нескольких сотен саркомеров. Тонкие нити присоединяются к сарколемме, толстые находятся внутри волокон. Ионы кальция также играют роль в сокращении, но поступают в мышцу не из цистерн, а из внеклеточного вещества, поскольку в гладких мышцах отсутствуют цистерны с ионами калькия. Этот процесс медленный и поэтому медленно работают гладкие мышцы.
Рисунок. Схема расположения толстых и тонких нией в гладких мышечных волокнах.
(одно занятие)
1. Назовите основные структурные элементы мышечного волокна, обеспечивающие его возбуждение и сокращение.
Клеточная мембрана (сарколемма), саркоплазматический ретикулум, миофибриллы.
2. Каково функциональное значение мембраны мышечного волокна в выполнении его сократительной функции?
Сарколемма является оболочкой для структурных элементов мышечного волокна, обеспечивает формирование потенциала покоя, возникновение потенциала действия и проведение возбуждения.
3. Что представляет собой миофибрилла, каково ее значение в механизме мышечного сокращения?
Миофибрилла – структура мышечного волокна, состоящая из большого числа протофибрилл (совокупность нитей актина и миозина);является сократительным элементом мышечного волокна.
4. Что представляет собой саркоплазматический ретикулум, каково его значение в механизме мышечного сокращения?
Это замкнутая система внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу. Является резервуаром для хранения, выброса и обратного захвата кальция при сокращении и расслаблении мышцы.
5. Назовите структурную и функциональную единицы изолированной мышцы и двигательного аппарата в организме. Что называют двигательной единицей?
Мышечное волокно и двигательная единица соответственно. Мотонейрон с группой иннервируемых им мышечных волокон.
6. На какие группы по скорости сокращения делятся двигательные единицы, какова продолжительность их сокращения?
На быстрые и медленные. 0,01 – 0,03 с и 0,1с, соответственно.
7. Назовите группы мышц, состоящие преимущественно из быстрых или медленных мышечных волокон.
Из быстрых – некоторые мышцы глаза, мышцы пальцев рук; из медленных – дыхательные мышцы, разгибатели конечностей и спины, обеспечивающие поддержание позы.
8. Назовите функциональные отличия быстрых и медленных двигательных единиц.
У быстрых двигательных единиц скорость и сила сокращения больше, но быстрее наступает утомление, у медленных – обратные взаимоотношения.
9. Перечислите свойства мышечной ткани.
Возбудимость, проводимость, сократимость, растяжимость, эластичность.
10. Перечислите основные функции скелетных мышц.
Обеспечивают все виды двигательной активности, поддержание определенной позы, дыхательную функцию, жевание, выработку тепла, способствуют движению крови и лимфы по сосудам к сердцу.
11. Что называют сократимостью мышцы? Что является непосредственной причиной сокращения (укорочения) мышцы?
Способность мышечной ткани изменять длину или напряжение. Скольжение нитей актина вдоль нитей миозина навстречу друг другу.
12. Почему потенциал действия считается инициатором мышечного сокращения? Дайте соответствующие пояснения.
Потенциал действия повышает проницаемость саркоплазматического ретикулума, что обеспечивает выход из него ионов кальция, необходимых для запуска процесса сокращения мышцы.
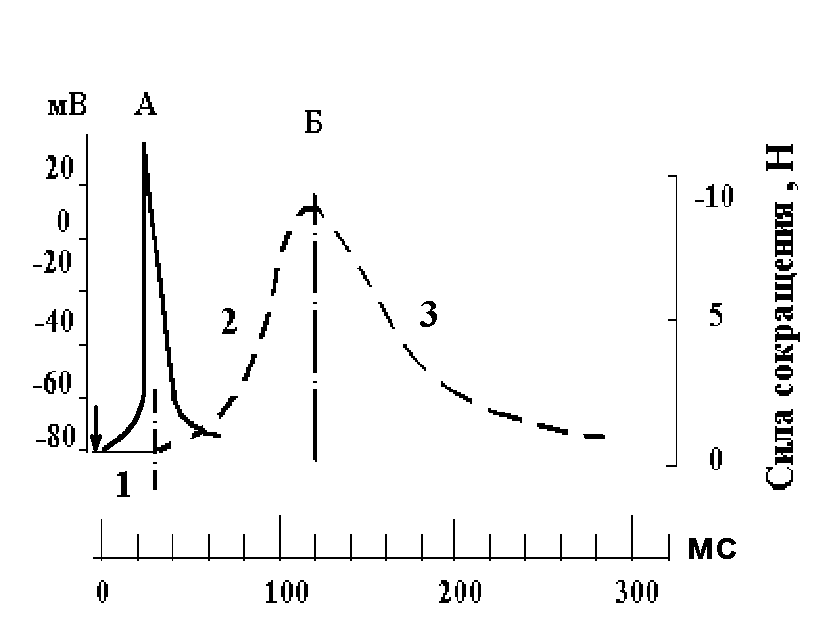
13. Нарисуйте потенциал действия скелетной мышцы, полученный при внутриклеточном отведении. Укажите его амплитуду в мВ.
14. Нарисуйте, сопоставив во времени, потенциал действия и цикл одиночного сокращения скелетной мышцы. Назовите фазы сокращения мышцы.
 А
– потенциал действия; Б – изометрическое
сокращение мышцы: 1 – латентный период;
2 – фаза напряжения; 3 – фаза расслабления;
– момент нанесения раздражения.
А
– потенциал действия; Б – изометрическое
сокращение мышцы: 1 – латентный период;
2 – фаза напряжения; 3 – фаза расслабления;
– момент нанесения раздражения.
15. Опишите кратко роль ионов кальция в механизме мышечного сокращения.
Ионы кальция взаимодействуют с белковым комплексом тропонин-тропомиозин, что ведет к освобождению активных участков на нитях актина и зацеплению за них головок миозиновых мостиков.
16. На какие процессы, обеспечивающие сокращение мышцы, расходуется энергия АТФ?
На взаимодействие актиновых и миозиновых нитей, обеспечивающее их скольжение относительно друг друга (укорочение) и работу ионных насосов.
17. Опишите последовательно процессы, обеспечивающие освобождение энергии АТФ при мышечном сокращении.
Контакт головок миозина с нитями актина – активация АТФ-азы миозина в присутствии ионов магния – расщепление АТФ – выделение энергии.
18. Что является непосредственной причиной скольжения нитей актина и миозина, обеспечивающего мышечное сокращение? Почему?
"Сгибание" миозиновых мостиков. Потому что в этот момент они «зацеплены» своими головками за активные участки нитей актина.
19. Активным (с затратой энергии АТФ) или пассивным (без затраты энергии АТФ) является процесс расслабления мышцы?
Расслабление мышцы обеспечивают как активные, так и пассивные процессы.
20. Какой из процессов, обеспечивающих мышечное расслабление, является активным, какой – пассивным?
Активным (с затратой энергии АТФ) является процесс переноса ионов кальция в саркоплазматический ретикулум, пассивным – скольжение нитей актина и миозина, ведущее к уменьшению зон их взаимного перекрытия.
21. Что является причиной скольжения нитей актина и миозина относительно друг друга при расслаблении мышцы.
Эластические свойства самой мышцы и сухожилия, растянутых при сокращении мышцы и масса органа (сила тяжести).
22. Назовите две фазы теплообразования в мышцах. Каким периодам мышечного сокращения соответствует первая из них?
Первая фаза – начальное теплообразование, соответствует возбуждению, укорочению и расслаблению мышцы, вторая – восстановительное (запаздывающее) теплообразование.
23. На какие процессы расходуется энергия восстановительного теплообразования в мышцах?
На ресинтез АТФ и работу ионных помп, обеспечивающих перенос ионов Na + , K + и Ca 2+ .
24. Назовите источники энергии, обеспечивающие ресинтез АТФ.
Расщепление креатинфосфата, анаэробный гликолиз; аэробное окисление углеводов и жирных кислот, т. е. через окислительное фосфорилирование.
25. На что расходуется энергия (укажите распределение в процентах), освобождаемая при мышечном сокращении? Каков КПД мышцы?
50% энергии расходуется на сокращение и расслабление (из них: 25% – на механическую деятельность [это КПД мышцы], 25% – на работу ионных помп); 50% тепла выделяется в окружающую среду.
26. Назовите типы сокращения скелетных мышц в зависимости от условий сокращения и от характера раздражения.
В зависимости от условий сокращения различают изометрическое и изотоническое сокращения. В зависимости от характера раздражения различают одиночное и тетаническое сокращения.
27. Назовите три фазы одиночного мышечного сокращения. Какой основной процесс происходит в первую фазу?
Латентный период, период укорочения и расслабления. Возбуждение.
28. Какие факторы влияют на силу одиночного мышечного сокращения?
Степень предварительного растяжения мышцы и сила ее раздражения.
29. Почему увеличение силы раздражения мышцы увеличивает силу ее сокращения?
Вследствие увеличения числа сокращающихся волокон. В возбуждение дополнительно вовлекаются волокна, которые при слабом раздражении не возбуждались из-за более низкой их возбудимости или более глубокого расположения в мышце.
30. Почему предварительное умеренное растяжение изолированной мышцы увеличивает силу ее сокращения при одиночном раздражении?
В результате того, что в умеренно растянутой мышце увеличивается как пассивное напряжение упругих элементов, так и активная сила сокращения вследствие увеличения зон взаимодействия нитей актина и миозина.
31. До какой степени необходимо растянуть изолированную мышцу, чтобы сила ее активного сокращения в изометрическом режиме была максимальной при одиночном раздражении? Почему?
До ее длины в состоянии покоя in situ, что обеспечивает максимальную зону контакта между нитями актина и миозина, и следовательно, максимальное число точек взаимодействия миозиновых мостиков с нитями актина.
32. Как и почему будет изменяться сила активного сокращения мышцы, если ее предварительно растянуть более ее длины в состоянии покоя in situ, а затем, перед каждым последующим раздражением увеличивать степень ее растяжения?
Будет уменьшаться вплоть до нуля в результате уменьшения зон взаимного перекрытия нитей актина и миозина (уменьшается число точек зацепления миозиновых мостиков с нитями актина, вплоть до полного их отсутствия).
33. Что называют тетаническим сокращением мышцы? Какое явление лежит в основе механизма тетануса?
Слитное, длительное сокращение скелетной мышцы, возникающее в ответ на ритмическое раздражение. Явление суммации мышечных сокращений.
34. Что называют суммацией мышечных сокращений?
Увеличение силы (или амплитуды) и длительности сокращения мышцы под действием ее повторного раздражения в период предыдущего сокращения.
35. За счет чего увеличивается амплитуда мышечного сокращения при суммации в изотоническом режиме? Объясните механизм.
За счет дополнительного скольжения нитей актина и миозина относительно друг друга в результате увеличения зон зацепления миозиновых мостиков под влиянием дополнительного выхода ионов Са 2+ из саркоплазматического ретикулума.
36. Почему при суммации мышечных сокращений в изометрическом режиме возможно дополнительное скольжения нитей актина относительно миозина?
Потому что мышца обладает растяжимостью, что делает возможным дополнительное укорочение саркомеров.
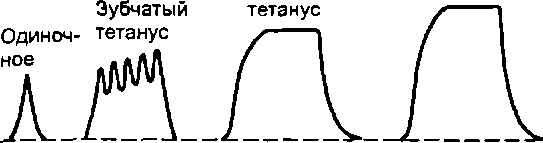
37. При каких условиях раздражения скелетной мышцы вместо одиночных сокращений возникает тетанус? Какие виды тетануса Вам известны?
При ритмическом раздражении с интервалами между импульсами меньшими, чем период одиночного мышечного сокращения. Зубчатый и гладкий.
38. В какую фазу одиночного сокращения должно попасть каждое последующее раздражение, чтобы возник зубчатый или гладкий тетанус? Какие факторы влияют на высоту гладкого тетануса изолированной мышцы?
В фазу расслабления мышцы или в фазу укорочения (напряжения) мышцы соответственно. Степень предварительного растяжения мышцы, сила и частота ее раздражения.
39. Какова зависимость высоты гладкого тетануса от частоты раздражения мышцы (в динамике)?
С увеличением частоты раздражения до известного предела величина тетанического сокращения нарастает, а затем уменьшается, вплоть до полного расслабления мышцы.
40. Какую частоту раздражения мышцы называют оптимальной, какую – пессимальной?
Оптимальная частота – при которой гладкий тетанус наиболее высокий и устойчивый, пессимальная – высокая частота, превышающая лабильность мышц, при которой она расслабляется.
41. Почему при оптимальной частоте раздражения изолированной мышцы тетанус наиболее высокий и устойчивый, а при пессимальной частоте раздражения мышца расслабляется?
Потому что при оптимальной частоте раздражения каждый последующий стимул попадает в фазу экзальтации, а при пессимальной – в фазу абсолютной рефрактерности.
42. В каком звене нервно-мышечного препарата (нерв-синапс-мышца) и почему при ритмическом раздражении нерва развивается пессимальное торможение Н. Е. Введенского?
В мионевральном синапсе, так как его лабильность наименьшая.
43. С помощью какого приема, можно доказать, что пессимум не связан с утомлением мионеврального синапса?
Уменьшение частоты раздражения до оптимальной сразу же ведет к восстановлению исходного гладкого тетануса.
44. Синхронно или асинхронно сокращаются отдельные мышечные волокна в естественных условиях? За счет каких механических эффектов увеличивается сила сокращения скелетной мышцы в естественных условиях?
Асинхронно. За счет вовлечения в реакцию большего числа двигательных единиц, увеличения степени синхронизации их возбуждения, дополнительного скольжения нитей актина и миозина относительно друг друга в каждой миофибрилле.
45. Подчиняется ли двигательная единица закону "все или ничего"? Почему?
Подчиняется, так как импульсы по ветвлениям аксона подходят одновременно ко всем мышечным волокнам двигательной единицы, и они сокращаются синхронно, т. е. двигательная единица функционирует как единое целое.
46. В каких условиях отдельные двигательные единицы одного нейронного пула возбуждаются синхронно, в каких – асинхронно?
При умеренных мышечных нагрузках – асинхронно, т. е. независимо друг от друга, при чрезмерных усилиях – синхронно.
47. Почему асинхронное возбуждение отдельных двигательных единиц в естественных условиях дает плавное тетаническое сокращение скелетных мышц?
Вследствие суммации сокращений большого числа асинхронно сокращающихся мышечных волокон различных двигательных единиц.
48. В каких отделах центральной нервной системы находятся мотонейроны, аксоны которых иннервируют скелетные мышцы?
В передних рогах спинного мозга, в продолговатом мозгу, в мосту и среднем мозге.
49. Что называют тонусом скелетных мышц, развивается ли при этом их утомление, велик ли расход энергии?
Постоянное слабое напряжение (сокращение) скелетных мышц, поддерживаемое редкими импульсами из центральной нервной системы и осуществляемое с малым расходом энергии без признаков утомления.
50. Какова зависимость работы изолированной скелетной мышцы от величины нагрузки?
С увеличением нагрузки работа мышцы сначала возрастает, а затем уменьшается вплоть до нуля при чрезмерно сильных нагрузках, когда мышца не в состоянии поднять груз.
51. Сформулируйте правило "средних нагрузок". Как и почему изменится работоспособность скелетной мышцы при увеличении частоты ее сокращений?
Работа мышцы максимальна при средних нагрузках. С увеличением частоты сокращений работоспособность сначала возрастает, а при превышении оптимальной частоты работоспособность уменьшается, т. к. быстрее развивается утомление.
52. Что называют утомлением мышцы? Чем оно объясняется?
Временное понижение работоспособности мышцы, наступающее в результате работы и исчезающее после отдыха. Накоплением продуктов обмена веществ, постепенным истощением энергетических запасов.
53. В условиях целого организма или в изолированной мышце утомление наступает медленнее? Почему?
В условиях целого организма, т. к. мышца постоянно снабжается кровью: получает питательные вещества и кислород и освобождается от продуктов метаболизма.
54. Где в условиях целого организма утомление наступает раньше: в центральной нервной системе, в нервно-мышечном синапсе или в самой мышце? Что такое активный отдых?
В центральной нервной системе. Восстановление работоспособности утомленных мышц при двигательной активности других мышц.
55. Как доказать в эксперименте, что в изолированной мышце утомление связано с накоплением продуктов обмена?
Изолированную мышцу поместить в раствор Рингера и длительным раздражением вызвать ее утомление, после замены раствора – работоспособность мышцы на некоторое время восстанавливается.
56. Перечислите структурные особенности гладкой мышцы.
Нерегулярное расположение нитей актина и миозина, вследствие чего отсутствует поперечная исчерченность, слабое развитие саркоплазматического ретикулума, наличие нексусов между мышечными волокнами.
57. Перечислите особенности потенциала покоя и потенциала действия гладкой мышцы по сравнению с таковыми поперечнополосатой мышцы.
Величина потенциала покоя гладкой мышцы меньше (30 – 50 мВ), наблюдается спонтанная деполяризация. Потенциалы действия бывают пикообразными и платообразными, более продолжительны – до 0, 5 с, деполяризация обеспечивается, главным образом, с помощью кальция и частично посредством натрия.
58. Назовите функциональные особенности гладкой мышцы по сравнению со скелетной.
Гладкой мышце присущи: автоматия, пластичность, более продолжительное сокращение (от нескольких секунд до 1 мин).
59. Что такое пластичность гладких мышц, каково ее значение для функционирования внутренних полых органов?
Способность длительно сохранять приданную растяжением длину без изменения напряжения. Благодаря пластичности гладкой мышцы давление в полых органах почти не меняется при медленном их наполнения.
60. Что является функциональной единицей гладкой мышцы? Почему?
Пучок мышечных волокон, заключенный в соединительнотканную оболочку, в пределах которого возбуждение передается от одного волокна к другому, охватывая весь пучок одновременно.
1. Какими свойствами обладает скелетная мышца плода к моменту рождения? Как изменяется упругость, прочность и эластичность мышц с возрастом?
Возбудимостью, проводимостью, сократимостью, эластичностью и растяжимостью, т. е. всеми свойствами мышцы взрослого. Упругость и прочность увеличиваются, эластичность уменьшается.
2. Как изменяются в онтогенезе возбудимость, проводимость, сократимость, утомляемость, быстрота сокращения и расслабления, лабильность скелетной мышцы?
Показатели всех перечисленных свойств увеличиваются.
3. Каковы соотношения силы мышц мальчиков и девочек в период от 7 до 8 лет, в возрасте 10 – 12 лет и 15 – 18 лет?
До 7 – 8 летнего возраста сила их мышц одинакова, в 10 – 12 лет – больше у девочек, в 15 – 18 лет – больше у мальчиков.
4. Укажите величину мембранного потенциала мышечного волокна новорожденного ребенка и взрослого человека. С чем связано это различие?
У новорожденных 20 – 40 мВ, у взрослых 80 –90 мВ. У новорожденных больше ионная проницаемость мембраны мышечного волокна, поэтому больше утечка ионов, меньше градиент их концентрации.
5. Перечислите отличия потенциала действия мышечного волокна новорожденного от такового у взрослого.
Меньшая амплитуда, большая продолжительность, часто отсутствует инверсия.
6. Увеличивается или уменьшается скорость проведения возбуждения по мышечному волокну с возрастом? Перечислите факторы, обеспечивающие это изменение.
Увеличивается. Увеличение мембранного потенциала, потенциала действия, а также диаметра мышечного волокна.
7. Почему увеличение амплитуды потенциала действия мышечного волокна в процессе роста организма увеличивает скорость проведения возбуждения?
Больший потенциал действия быстрее вызывает возбуждение соседнего участка мышечного волокна.
8. Почему увеличение диаметра мышечного волокна в процессе роста организма увеличивает скорость проведения возбуждения?
Увеличение толщины мышечного волокна ведет к уменьшению продольного сопротивления ионному току в миоплазме.
9. Перечислите особенности сокращения мышц новорожденного. Разделяются ли мышцы новорожденного на быстрые и медленные?
Относительная длительность одиночного сокращения (фазы укорочения и расслабления), тонические сокращения, без признаков пессимального торможения при большой частоте раздражения. Не подразделяются.
10. Как изменяется эффективность отдыха (становится больше или меньше) после физического утомления у детей разного возраста: 7 – 12 лет, 13 – 15 лет и в 16 – 18 лет?
В 7 – 12 лет эффективность отдыха наибольшая, в 13 – 15 лет резко падает, в 16 – 18 лет несколько увеличивается.
11. В каком возрасте наблюдается максимальная выносливость к физическим нагрузкам?
В возрасте 20 – 29 лет.
12. Что представляет собой незрелый (примитивный) нервно-мышечный синапс новорожденного? В чем заключается его функциональная особенность, к какому возрасту заканчивается его созревание?
На поверхности миотрубки выемка, в которой расположено окончание аксона. Значительно большая синаптическая задержка (в 7 – 10 раз больше, чем у взрослых). К 7 – 8 годам.
13. В чем выражается созревание терминальных ветвлений аксона мотонейрона? Какое значение имеет этот процесс?
В увеличении терминальных ветвлений аксона и содержания в них ацетилхолина. Это ведет к увеличению выхода медиатора в синаптическую щель при поступлении импульса к нервному окончанию и увеличению амплитуд ПКП.
14. В чем выражается созревание постсинаптической мембраны?
В увеличении плотности холинорецепторов на ней, образовании складок, увеличении потенциала концевой пластинки, появлении в ней холинэстеразы.
15. Как и почему изменяется синаптическая задержка в нервно-мышечном синапсе в процессе созревания?
Уменьшается вследствие увеличения скорости освобождения ацетилхолина из пресинаптического окончания на каждый нервный импульс, увеличения потенциала концевой пластинки и появления в ней холинэстеразы.
Скелетные мышцы - активная часть опорно-двигательного аппарата, включающего также кости, связки, сухожилия и их сочленения. С функциональной точки зрения к двигательному аппарату можно отнести и мотонейроны, вызывающие возбуждение мышечных волокон. Аксон мотонейрона при входе в скелетную мышцу ветвится, и каждая веточка участвует в формировании нервно-мышечного синапса на отдельном мышечном волокне.
Мотонейрон вместе с иннервируемыми им мышечными волокнами называют нейромоторной (или двигательной) единицей (ДЕ). В глазных мышцах одна двигательная единица содержит 13-20 мышечных волокон, в мышцах туловища - со 1 тни волокон, в камбаловидной мышце - 1500-2500 волокон. Мышечные волокна одной ДЕ имеют одинаковые морфофунк- циональные свойства.
Функциями скелетных мышц являются: 1) передвижение тела в пространстве; 2) перемещение частей тела относительно друг друга, втом числе осуществление дыхательных движений, обеспечивающих вентиляцию легких; 3) поддержание положения и позы тела. Кроме того, поперечно-полосатые мышцы имеют значение в выработке тепла, поддерживающего температурный гомеостаз, и в депонировании некоторых питательных веществ.
Физиологические свойства скелетных мышц выделяют:
1) возбудимость. Из-за высокой поляризации мембран поперечно-полосатых мышечных волокон (90 мВ) возбудимость их ниже, чем у нервных волокон. Амплитуда потенциала действия у них (130 мВ) больше, чем удругих возбудимых клеток. Это позволяет на практике достаточно легко регистрировать биоэлектрическую активность скелетных мышц. Длительность потенциала действия составляет 3-5 мс. Этим определяется короткий период абсолютной рефрактерности мышечных волокон;
проводимость. Скорость проведения возбуждения вдоль мембраны мышечного волокна составляет 3-5 м/с;
сократимость. Представляет специфическое свойство мышечных волокон изменять свою длину и напряжение при развитии возбуждения.
Скелетные мышцы обладают также эластичностью и вязкостью.
Режимы и виды мышечных сокращений. Изотонический режим - мышца укорачивается при отсутствии возрастания ее напряжения. Такое сокращение возможно только для изолированной (удаленной из организма) мышцы.
Изометрический режим - напряжение мышцы возрастает, а длина практически не уменьшается. Такое сокращение наблюдается при попытке поднять непосильный груз.
Ауксотонический режим мышца укорачивается и возрастает ее напряжение. Такое сокращение чаще всего наблюдается при осуществлении трудовой деятельности человека. Вместо термина "ауксотонический режим" часто применяется название концентрический режим.
Выделяют два вида мышечных сокращений: одиночное и те- таническое.
Одиночное мышечное сокращение проявляется в результате развития одиночной волны возбуждения в мышечных волокнах. Этого можно достичь при воздействии на мышцу очень коротким (около 1 мс) стимулом. В развитии одиночного мышечного сокращения выделяют латентный период, фазу укорочения и фазу расслабления. Сокращение мышцы начинает проявляться через 10 мс от начала воздействия раздражителя. Этот временной интервал называют латентным периодом (рис.5.1). Затем последует развитие укорочения (длительность около 50 мс) и расслабления (50-60 мс). Считается, что на весь цикл одиночного мышечного сокращения затрачивается в среднем 0,1 с. Но следует иметь в виду, что длительность одиночного сокращения у разных мышц может сильно варьировать. Она также зависит от функционального состояния мышцы. Скорость сокращения и особенно расслабления замедляется при развитии утомления мышцы. К быстрым мышцам, имеющим короткий период одиночного сокращения, относятся мышцы языка и смыкающие веко.
Рис. 5.1. Временные соотношения разных проявлений возбуждения волокна скелетной мышцы: а - соотношение потенциала действия, выхода Са 2+ в саркоплазму и сокращения: / - латентный период; 2 - укорочение; 3 - расслабление; б - соотношение потенциала действия, сокращения и уровня возбудимости
Под влиянием одиночного раздражителя вначале возникает потенциал действия и лишь затем начинает развиваться период укорочения. Оно продолжается и после окончания реполяризации. Восстановление исходной поляризации сарколеммы свидетельствует и о восстановлении возбудимости. Следовательно, на фоне развивающегося сокращения в мышечных волокнах можно вызвать новые волны возбуждения, сократительный эффект от которых будет суммироваться.
Тетаническим сокращением или тетанусом называют сокращение мышцы, появляющееся в результате возникновения в моторных единицах многочисленных волн возбуждения, сократительный эффект от которых суммируется по амплитуде и времени.
Различают зубчатый и гладкий тетанус. Для получения зубчатого тетануса надо стимулировать мышцу с такой частотой, чтобы каждое последующее воздействие наносилось после фазы укорочения, но до момента окончания расслабления. Гладкий тетанус получается при более частых раздражениях, когда последующие воздействия наносятся во время развития укорочения мышцы. Например, если фаза укорочения у мышцы составляет 50 мс, а фаза расслабления - 60 мс, то для получения зубчатого тетануса необходимо раздражать эту мышцу с частотой 9-19 Гц, для получения гладкого - с частотой не менее 20 Гц.
Несмотря
Амплитуда сокращений
расслабилась
Пессимум
на длящееся раздражение, мышца
30 Гц
1 Гц 7 Гц
200 Гц
50 Гц
Частота раздраженияРис. 5.2. Зависимость амплитуды сокращения от частоты раздражения (сила и длительность стимулов неизменны)
Для демонстрации различных видов тетануса обычно используют регистрацию сокращений изолированной икроножной мышцы лягушки на кимографе. Пример такой кимограм- мы представлен на рис. 5.2. Амплитуда одиночного сокращения минимальна, увеличивается при зубчатом тетанусе и становится максимальной - при гладком. Одной из причин такого возрастания амплитуды является то, что при возникновении частых волн возбуждения в саркоплазме мышечных волокон накапливается Са 2+ стимулирующий взаимодействие сократительных белков.
При постепенном увеличении частоты раздражения нарастание силы и амплитуды сокращения мышцы идет лишь до некоторого предела - оптимума ответной реакции. Частоту раздражения, вызывающую наибольший ответ мышцы, называют оптимальной. Дальнейшее увеличение частоты раздражения сопровождается уменьшением амплитуды и силы сокращения. Это явление называют пессимумом ответной реакции, а частоты раздражения, превышающие оптимальную величину, - пессимальными. Явления оптимума и пессимума были открыты Н.Е. Введенским.
При оценке функциональной активности мышц говорят об их тонусе и фазических сокращениях. Тонусом мышцы называют состояние длительного непрерывного напряжения. При этом видимое укорочение мышцы может отсутствовать из-за того, что возбуждение возникает не во всех, а лишь в некоторых моторных единицах мышцы и они возбуждаются не синхронно. Фазическим сокращением мышцы называют кратковременное укорочение мышцы, сменяющееся ее расслаблением.
Структурно -функциональная характеристика мышечного волокна. Структурной и функциональной единицей скелетной мышцы является мышечное волокно, представляющее собой вытянутую (длиной 0,5-40 см) многоядерную клетку. Толщина мышечных волокон составляет 10- 100 мкм. Диаметр их может увеличиваться при интенсивных тренировочных нагрузках, количество же мышечных волокон может нарастать лишь до 3-4-месячного возраста.
Мембрану мышечного волокна называют сарколеммой, цитоплазму - саркоплазмой. В саркоплазме располагаются ядра, многочисленные органеллы, саркоплазматический рети- кулум, в состав которого входят продольные трубочки и их утолщения - цистерны, в которых содержатся запасы Са 2+ Цистерны соседствуют с поперечными трубочками, пронизывающими волокно в поперечном направлении (рис. 5.3).
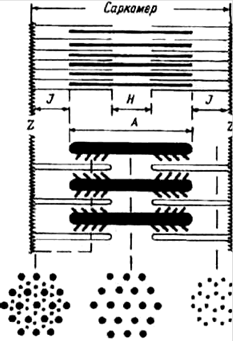
В саркоплазме вдоль мышечного волокна проходит около 2000 миофибрилл (толщиной около 1 мкм), которые включают нити, образованные сплетением молекул сократительных белков: актина и миозина. Молекулы актина образуют тонкие нити (миофиламенты), которые лежат параллельно друг другу и пронизывают своеобразную мембрану, называемую Z-линией или полоской. Z-линии расположены перпендикулярно длинной оси миофибриллы и делят миофибриллу на участки длиной 2- 3 мкм. Эти участки называют саркомерами.
Цистерна Сарколемма
Поперечная трубочка
Саркомер
Трубочка с-п. рет^|Jj3H сссс с_ з зззз tccc ;
; зззз сссс с
з зззз сссс с
j3333 СССС£
J3333 с с с с с_
J3333 сс с с с_
Саркомер сокращен
3 3333 сссс сСаркомер расслаблен
Рис. 5.3. Строение саркомера мышечного волокна: Z-линии - ограничивают саркомер,/! - анизотропный (темный)диск, / - изотропный (светлый) диск, Н - зона (менее темная)
Саркомер является сократительной единицей миофибрил- лы- В центре саркомера строго упорядоченно друг над другом лежат толстые нити, сформированные молекулами миозина, flo краям саркомера аналогичным образом расположены тонкие нити актина. Концы актиновых нитей заходят между концами миозиновых нитей.
Центральная часть саркомера (ширина 1,6 мкм), в которой лежат мио- зиновые нити, под микроскопом выглядит темной. Этот темный участок прослеживается поперек всего мышечного волокна, так как саркомеры соседних миофибрилл располагаются строго симметрично друг над другом. Темные участки саркомеров получили название А-дисков от слова "анизотропный" Эти участки обладают двойным лучепреломлением в поляризованном свете. Зоны по краям А-диска, где нити актина и миозина перекрываются, кажутся темнее, чем в центре, где находятся только миозиновые нити. Этот центральный участок называют полоской Н.
Участки миофибриллы, в которых располагаются только актиновые нити, не обладают двойным лучепреломлением, они изотропны. Отсюда их название - I-диски. В центре I-диска проходит узкая темная линия, образованная Z-мембраной. Эта мембрана удерживает в упорядоченном состоянии актиновые нити двух соседних саркомеров.
В состав актиновой нити кроме молекул актина входят также белки тропомиозин и тропонин, влияющие на взаимодействие нитей актина и миозина. В молекуле миозина выделяют участки, которые называют головкой, шейкой и хвостом. В каждой такой молекуле имеется один хвост и по две головки с шейками. На каждой головке имеется химический центр, который может присоединять АТФ и участок, позволяющий связываться с актиновой нитью.
Молекулы миозина при формировании миозиновой нити сплетаются своими длинными хвостами, располагающимися в центре этой нити, а головки находятся ближе к ее концам (рис. 5.4). Шейка и головка образуют выступ, торчащий из миозиновых нитей. Эти выступы называют поперечными мостиками. Они подвижны, и благодаря таким мостикам миозиновые нити могут установить связь с актиновыми.
Когда к головке молекулы миозина присоединяется АТФ, то мостик на короткое время располагается под тупым углом относительно хвоста. В следующий момент происходит частичное расщепление АТФ и за счет этого головка приподнимается, переходит в энергизированное положение, при котором она может связываться с актиновой нитью.
Молекулы актина образуют двойную спираль Тролонин
Центр связи с АТФ
Участок тонкой нити (вдоль цепочек актина располагаются молекулы тропомиозина, тролонин в узлах спирали)
Шейка
Хвост
Тропомиоэин т iМолекула миозина при большом увеличении
Участок толстой нити (видны головки молекул миозина)
Нить актина
Головка

+Са 2+
Са 2+ "*Са 2+
АДФ- Ф
Са 2+ N
Расслабление
Цикл движений головки миозина при сокращении мышцы
миозина 0 +АТФРис. 5.4. Структура нитей актина и миозина, движение головок миозина при сокращении и расслаблении мышцы. Объяснение в тексте: 1-4 - этапы цикла
Механизм сокращения мышечного волокна. Возбуждение волокна скелетной мышцы в условиях физиологической нормы вызывается только импульсами, приходящими от мотонейронов. Нервный импульс активирует нервно-мышечный синапс, вызывает возникновение ПК.П, а потенциал концевой пластинки обеспечивает генерацию потенциала действия на сарколемме.
Потенциал действия распространяется как вдоль поверхностной мембраны мышечного волокна, так и вглубь по поперечным трубочкам. При этом происходит деполяризация цистерн саркоплазматического ретикулума и открытие Са 2+ -каналов. Поскольку в саркоплазме концентрация Са 2+ составляет 1(Г 7 -1(Г б М, а в цистернах она приблизительно в 10 ООО раз большая, то при открытии Са 2+ -каналов кальций по градиенту концентрации выходит из цистерн в саркоплазму, диффундирует к миофиламентам и запускает процессы, обеспечивающие сокращение. Таким образом, выход ионов Са 2+
в саркоплазму является фактором, сопрягающим электрические и механические явления в мышечном волокне. Ионы Са 2+ связываются с тропонином и это, при участии тропомио- зина, приводит к открытию (разблокировке) участков актиновой нити, которые могут связываться с миозином. После этого энергизированные головки миозина образуют мостики с актином, происходит окончательное расщепление АТФ, ранее захваченных и удерживаемых головками миозина. Получаемая от расщепления АТФ энергия идет на поворот головок миозина в направлении к центру саркомера. При таком повороте головки миозина тянут за собой актиновые нити, продвигая их между миозиновыми. За одно грёбковое движение головка может продвинуть актиновую нить на-1 % от длины саркомера. Для максимального сокращения нужны повторные гребковые движения головок. Это имеет место при достаточной концентрации АТФ и Са 2+ в саркоплазме. Для повторного движения головки миозина необходимо, чтобы к ней присоединилась новая молекула АТФ. Подсоединение АТФ вызывает разрыв связи головки миозина с актином и она на мгновение занимает исходное положение, из которого может переходить к взаимодействию с новым участком актиновой нити и делать новое гребковое движение.
Такую теорию механизма мышечного сокращения назвали теорией "скользящих нитей"
Для расслабления мышечного волокна необходимо, чтобы концентрация ионов Са 2+ в саркоплазме стала менее Ю -7 М/л. Это происходит за счет функционирования кальциевого насоса, который перегоняет Са 2+ из саркоплазмы в ретикулум. Кроме того, для расслабления мышцы необходимо, чтобы были разорваны мостики между головками миозина и актином. Такой разрыв происходит при наличии в саркоплазме молекул АТФ и связывания их с головками миозина. После отсоединения головок эластические силы растягивают саркомер и перемещают нити актина в исходное положение. Эластические силы формируются за счет: 1) эластической тяги спиралевидных клеточных белков, входящих в структуру саркомера; 2) эластических свойств мембран саркоплазматического ретикулума и сарколеммы; 3) эластичности соединительной ткани мышцы, сухожилий и действия сил гравитации.
Сила мышц. Силу мышцы определяют по максимальной Величине груза, который она может поднять, либо по максимальной силе (напряжению), которую она может развить в условиях изометрического сокращения.
Одиночное мышечное волокно способно развить напряжение 100-200 мг. В теле примерно 15-30 млн волокон. Если бы они действовали параллельно в одном направлении и одновременно, то могли бы создать напряжение 20-30 т.
Сила мышц зависит от ряда морфофункциональных, физиологических и физических факторов.
Сила мышц возрастает с увеличением площади их геометрического и физиологического поперечного сечения. Для определения физиологического поперечного сечения мышцы находят сумму поперечных сечений всех волокон мышцы по линии, проведенной перпендикулярно к ходу каждого мышечного волокна.
В мышце с параллельным ходом волокон (портняжная) геометрическое и физиологическое поперечные сечения равны. В мышцах с косым ходом волокон (межреберные) физиологическое сечение больше геометрического и это способствует увеличению силы мышц. Еще больше возрастает физиологическое сечение и сила у мышц с перистым расположением (большинство мышц тела) мышечных волокон.
Чтобы иметь возможность сопоставить силу мышечных волокон в мышцах с различным гистологическим строением, ввели понятие абсолютной силы мышцы.
Абсолютная сила мышцы - максимальная сила, развиваемая мышцей, в перерасчете на 1 см 2 физиологического поперечного сечения. Абсолютная сила бицепса - 11,9 кг/см 2 , трехглавой мышцы плеча - 16,8 кг/см 2 , икроножной 5,9 кг/см 2 , гладкой - 1 кг/см 2
Сила мышцы зависит от процентного соотношения различных типов двигательных единиц, входящих в эту мышцу. Соотношение разных типов двигательных единиц в одной и той же мышце у людей неодинаково.
Выделяют следующие типы двигательных единиц: а) медленные, неутомляемые (имеют красный цвет) - обладают малой силой, но могут быть длительно в состоянии тонического сокращения без признаков утомления; б) быстрые, легко- утомляемые (имеют белый цвет) - их волокна обладают большой силой сокращения; в) быстрые, устойчивые к утомлению - имеют относительно большую силу сокращения и в них медленно развивается утомление.
У разных людей соотношение числа медленных и быстрых двигательных единиц в одной и той же мышце определено генетически и может значительно различаться. Так, в четырехглавой мышце бедра человека относительное содержание мед- денных волокон может варьировать от 40 до 98%. Чем больший процент медленных волокон в мышцах человека, тем более они приспособлены к длительной, но небольшой по мощности работе. Люди с высоким содержанием быстрых сильных моторных единиц способны развивать большую силу, но склонны к быстрому утомлению. Однако надо иметь в виду, что утомление зависит и от многих других факторов.
Сила мышцы увеличивается при умеренном ее растяжении. Это происходит из-за того, что при умеренном растяжении саркомера (до 2,2 мкм) увеличивается количество мостиков, которые могут образоваться между актином и миозином. При растяжении мышцы в ней также развивается эластическая тяга, направленная на укорочение. Эта тяга суммируется с силой, развиваемой движением головок миозина.
Сила мышц регулируется нервной системой путем изменения частоты импульсаций, посылаемых к мышце, синхронизации возбуждения большого числа моторных единиц, выбора типов моторных единиц. Сила сокращений увеличивается: а) при возрастании количества возбуждаемых моторных единиц, вовлекаемых в ответную реакцию; б) при увеличении частоты волн возбуждения в каждом из активируемых волокон; в) при синхронизации волн возбуждения в мышечных волокнах; г) при активации сильных (белых) моторных единиц.
Сначала (при необходимости развития небольшого усилия) активируются медленные неутомляемые моторные единицы, затем быстрые, устойчивые к утомлению. А если надо развить силу более 20-25% от максимальной, то в сокращение вовлекаются быстрые легкоутомляемые моторные единицы.
При напряжении до 75% от максимально возможного практически все моторные единицы активированы и дальнейший прирост силы идет за счет увеличения частоты импульсов, приходящих к мышечным волокнам.
При слабых сокращениях частота импульсаций в аксонах мотонейронов составляет 5-10 имп/с, а при большой силе сокращения может доходить до 50 имп/с.
В детском возрасте прирост силы идет главным образом за счет увеличения толщины мышечных волокон, и это связано с увеличением количества миофибрилл. Увеличение числа волокон незначительно.
При тренировке мышцу взрослых нарастание их силы связано с увеличением числа миофибрилл, повышение же выносливости обусловлено увеличением числа митохондрий и интенсивности синтеза АТФ за счет аэробных процессов.
Существует взаимосвязь силы и скорости укорочения. Скорость сокращения мышцы тем выше, чем больше ее длина (за счет суммации сократительных эффектов саркомеров) и зависит от нагрузки на мышцу. При увеличении нагрузки скорость сокращения уменьшается. Тяжелый груз можно поднять только при медленном движении. Максимальная скорость сокращения, достигаемая при сокращении мышц человека, около 8 м/с.
Сила сокращения мышцы снижается при развитии утомления.
Утомление и его физиологические основы. Утомлением называют временное понижение работоспособности, обусловленное предыдущей работой и исчезающее после периода отдыха.
Утомление проявляется снижением мышечной силы, скорости и точности движений, изменением показателей работы кардиореспираторной системы и вегетативных регуляций, ухудшением показателей функций центральной нервной системы. О последнем свидетельствует снижение скорости простейших психических реакций, ослабление внимания, памяти, ухудшение показателей мышления, возрастание количества ошибочных действий.
Субъективно утомление может проявляться ощущением усталости, появлением боли в мышцах, сердцебиением, симптомами одышки, желанием снизить нагрузку или прекратить работу. Симптомы усталости могут различаться в зависимости от вида работы, ее интенсивности и степени утомления. Если утомление вызвано умственной работой, то, как правило, более выражены симптомы снижения функциональных возможностей психической деятельности. При очень тяжелой мышечной работе на первый план могут выступать симптомы нарушений на уровне нервно-мышечного аппарата.
Утомление, развивающееся в условиях обычной трудовой деятельности как при мышечной, так и при умственной работе, имеет во многом сходные механизмы развития. В обоих случаях процессы утомления раньше всего развиваются в нервных центрах. Одним из показателей этого является снижение умственной работоспособности при физическом утомлении, а при умственном утомлении - снижение эффективности мышечной деятельности.
Отдыхом называют состояние покоя или выполнение новой деятельности, при которых устраняется утомление и восстанавливается работоспособность. И.М. Сеченов показал, что восстановление работоспособности происходит быстрее, если при отдыхе после утомления одной группы мышц(напри- мер, левой руки), выполнять работу другой группой мышц (правой рукой). Это явление он назвал "активным отдыхом"
Восстановлением называют процессы, обеспечивающие ликвидацию дефицита запасов энергетических и пластических веществ, воспроизведение израсходованных или поврежденных при работе структур, устранение избытка метаболитов и отклонений показателей гомеостаза от оптимального уровня.
Длительность периода, необходимого для восстановления организма, зависит от интенсивности и длительности работы. Чем больше интенсивность труда, тем через более короткое время необходимо делать периоды отдыха.
Различные показатели физиологических и биохимических процессов восстанавливаются через разное время от момента окончания физической нагрузки. Одним из важных тестов скорости восстановления является определение времени, в течение которого частота сердечных сокращений возвращается к уровню, характерному для периода покоя. Время восстановления частоты сердечных сокращений после теста с умеренной физической нагрузкой у здорового человека не должно превышать 5 мин.
При очень интенсивной физической нагрузке явления утомления развиваются не только в центральной нервной системе, но и в нервно-мышечных синапсах, а также мышцах. В системе нервно-мышечного препарата наименьшей утомляемостью обладают нервные волокна, наибольшей - нервно- мышечный синапс, промежуточное положение занимает мыш- Ца. Нервные волокна часами могут проводить высокую частоту потенциалов действия без признаков утомления. При частой Же активации синапса эффективность передачи возбуждения сначала уменьшается, а затем наступает блокада его проведения. Это происходит из-за снижения запаса медиатора и АТФ в пресинаптической терминали, снижения чувствительности постсинаптической мембраны к ацетилхолину.
Был предложен ряд теорий механизма развития утомления в очень интенсивно работающей мышце: а) теория "истощения" - израсходование запасов АТФ и источников ее образования (креатинфосфата, гликогена, жирных кислот), б)теория "удушения" - на первое место выдвигается недостаток доставки кислорода в волокна работающей мышцы; в) теория "засорения", объясняющая утомление накоплением в мышце молочной кислоты и токсичных продуктов обмена веществ. В настоящее время считается, что все эти явления имеют место при очень интенсивной работе мышцы.
Установлено, что максимальная физическая работа до развития утомления выполняется при средней тяжести и темпе труда (правило средних нагрузок). В профилактике утомления важны также: правильное соотношение периодов труда и отдыха, чередование умственной и физической работы, учет околосуточных (циркадных), годовых и индивидуальных биологических ритмов.
Мощность мышцы равна произведению мышечной силы на скорость укорочения. Максимальная мощность развивается при средней скорости укорочения мышц. Для мышцы руки максимальная мощность (200 Вт) достигается при скорости сокращения 2,5 м/с.
5.2. Гладкие мышцы
Физиологические свойства и особенности гладких мышц.
Гладкие мышцы являются составной частью некоторых внутренних органов и участвуют в обеспечении функций, выполняемых этими органами. В частности, регулируют проходимость бронхов для воздуха, кровотока в различных органах и тканях, перемещения жидкостей и химуса (в желудке, кишечнике, мочеточниках, мочевом и желчном пузырях), осуществляют изгнание плода из матки, расширяют или сужают зрачки (за счет сокращения радиальных или циркулярных мышц радужной оболочки), изменяют положение волос и кожного рельефа. Гладкомышечные клетки имеют веретенообразную форму, длину 50-400 мкм, толщину 2-10 мкм.
Гладкие мышцы, как и скелетные, обладают возбудимостью, проводимостью и сократимостью. В отличие от скелетных м ышц, имеющих эластичность, гладкие - пластичны (способны длительное время сохранять приданную им за счет растяжения длину без увеличения напряжения). Такое свойство важно для выполнения функции депонирования пищи в желудке или жидкостей в желчном и мочевом пузырях.
Особенности возбудимости гладкомышечных волокон в определенной мере связаны с их низким трансмембранным потенциалом (Е 0 = 30-70 мВ). Многие из этих волокон обладают автоматией. Длительность потенциала действия у них может достигать десятков миллисекунд. Так происходит потому, что потенциал действия в этих волокнах развивается преимущественно за счет входа кальция в саркоплазму из межклеточной жидкости через так называемые медленные Са 2+ -каналы.
Скорость проведения возбуждения в гладкомышечных клетках малая - 2-10 см/с. В отличие от скелетных мышц возбуждение в гладкой мышце может передаваться с одного волокна на другое, рядом лежащее. Такая передача происходит благодаря наличию между гладкомышечными волокнами нексусов, обладающих малым сопротивлением электрическому току и обеспечивающих обмен между клетками Са 2+ и другими молекулами. В результате этого гладкая мышца имеет свойства функционального синтиция.
Сократимость гладкомышечных волокон отличается продолжительным латентным периодом (0,25-1,00 с) и большой длительностью (до 1 мин) одиночного сокращения. Гладкие мышцы имеют малую силу сокращения, но способны длительно находиться в тоническом сокращении без развития утомления. Это связано с тем, что на поддержание тетанического сокращения гладкая мышца расходует в 100-500 раз меньше энергии, чем скелетная. Поэтому расходуемые гладкой мышцей запасы АТФ успевают восстанавливаться даже во время сокращения и гладкие мышцы некоторых структур организма всю жизнь находятся в состоянии тонического сокращения.
Условия сокращения гладкой мышцы. Важнейшей особенностью гладкомышечных волокон является то, что они возбуждаются под влиянием многочисленных раздражителей. Сокращение скелетной мышцы в норме инициируется только нервным импульсом, приходящим к нервно-мышечному синапсу. Сокращение гладкой мышцы может быть вызвано как нервными Импульсами, так и биологически активными веществами (гормонами, многими нейромедиаторами, простагландинами, некоторыми метаболитами), а также воздействием физических факторов, например растяжением. Кроме того, возбуждение гладкой мышцы может произойти спонтанно - за счет автоматии.
Очень высокая реактивность гладких мышц, их свойство отвечать сокращением на действие разнообразных факторов создают значительные трудности для коррекции нарушений тонуса этих мышц в медицинской практике. Это видно на примерах лечения бронхиальной астмы, артериальной гипертонии, спастического колита и других заболеваний, требующих коррекции сократительной активности гладких мышц.
В молекулярном механизме сокращения гладкой мышцы также имеется ряд отличий от механизма сокращения скелетной мышцы. Нити актина и миозина в гладкомышечных волокнах располагаются менее упорядоченно, чем в скелетных, и поэтому гладкая мышца не имеет поперечной исчерченности. В актино- вых нитях гладкой мышцы нет белка тропонина и молекулярные центры актина всегда открыты для взаимодействия с головками миозина. Чтобы такое взаимодействие произошло, необходимо расщепление молекул АТФ и перенос фосфата на головки миозина. Тогда молекулы миозина сплетаются в нити и связываются своими головками с миозином. Далее следует поворот головок миозина, при котором актиновые нити втягиваются между миозиновыми и происходит сокращение.
Фосфорилирование головок миозина производится с помощью фермента киназы легких цепей миозина, а дефосфорили- рование - фосфатазы легких цепей миозина. Если активность фосфатазы миозина преобладает над активностью киназы, то головки миозина дефосфорилируются, связь миозина и актина разрывается и мышца расслабляется.
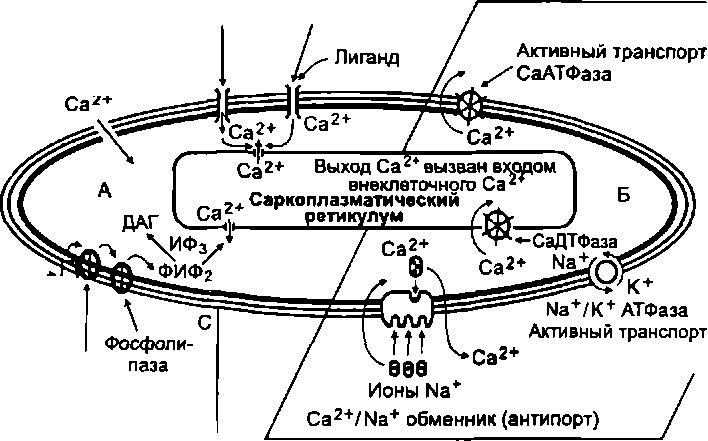
Следовательно, чтобы произошло сокращение гладкой мышцы, необходимо повышение активности киназы легких цепей миозина. Ее активность регулируется уровнем Са 2+ в саркоплазме. При возбуждении гладкомышечного волокна содержание кальция в его саркоплазме увеличивается. Это увеличение обусловлено поступлением Са^ + из двух источников: 1) межклеточного пространства; 2) саркоплазматического ре- тикулума (рис. 5.5). Далее ионы Са 2+ образуют комплекс с белком кальмодулином, который переводит в активное состояние киназу миозина.
Последовательность процессов, приводящих к развитию сокращения гладкой мышцы: вход Са 2 в саркоплазму - акти
вация кальмодулина (путем образования комплекса 4Са 2+ - кальмодулин) - активация киназы легких цепей миозина - фосфорилирование головок миозина - связывание головок миозина с актином и поворот головок, при котором нити актина втягиваются между нитями миозина.
Условия, необходимые для расслабления гладкой мышцы: 1) снижение (до 10 М/л и менее) содержания Са 2+ в саркоплазме; 2) распад комплекса 4Са 2+ -кальмодулин, приводящий к снижению активности киназы легких цепей миозина - дефосфорилирование головок миозина, приводящее к разрыву связей нитей актина и миозина. После этого силы упругости вызывают относительно медленное восстановление исходной длины гладкомышечного волокна, его расслабление.
Контрольные вопросы и задания
Клеточная мембрана
Рис. 5.5. Схема путей поступления Са 2+ в саркоплазму гладкомышеч-
ной клетки и удаления его из плазмы: а - механизмы, обеспечивающие поступление Са 2 + в саркоплазму и запуск со- кращеня (Са 2+ поступает из внеклеточной среды и саркоплазматического рети- кулума); б - пути удаления Са 2+ из саркоплазмы и обеспечения расслабления
Влияние норадреналина через а-адренорецепторы
Лигандзависимый Са 2+ -канал
Каналы "утечки г
Потенциал зависимый Са 2+ -канал
Гладкомышечная клетка
а-адрено! рецептор f Норадре- налин G
Назовите виды мышц человека. Каковы функции скелетных мышц?Дайте характеристику физиологических свойств скелетных мышц.
Каково соотношение потенциала действия, сокращения и возбудимости мышечного волокна?
Какие существуют режимы и виды мышечных сокращений?
Дайте структурно-функциональную характеристику мышечного волокна.
Что такое моторные единицы? Перечислите их виды и особенности.
Каков механизм сокращения и расслабления мышечного волокна?
Что такое сила мышц и какие факторы на нее влияют?
Какова связь между силой сокращения, его скоростью и работой?
Дайте определение утомления и восстановления. Каковы их физиологические основы?
Каковыфизиологические свойства и особенности гладких мышц?
Перечислите условия сокращения и расслабления гладкой мышцы.
Министерство здравоохранения Российской федерации
Казанский государственный медицинский университет
Кафедра физиологии
Механизм сокращения скелетной мышцы. Роль ионов кальция. Расслабление скелетной мышцы. Значение АТФ.
Контрольная
работа
(реферат)
студента гр.5102
Белокурова
Надежда Витальевна
Руководитель:__________ ______
_______________________ _______
Казань-2012г
- Введение.
Скелетная мускулатура человека и позвоночных животных состоит из мышечных волокон нескольких типов, отличающихся друг от друга стрктурно- функциональными характеристиками. В настоящие время выделяют четыре основных типа мышечных волокон.
Медленные физические волокна окислительного типа. Волокна этого типа характеризуются большим содержанием белка миоглобина, который способен связывать О 2 (близок по своим свойствам к гемоглобину). Мышцы, которые преимущественно состоят из волокон этого типа, за их красный тип называют красными. Они выполняют очень важную функцию поддержания позы человека и животных. Предельное утомление этих волокон, а следовательно и мышц, наступает очень медленно, что обусловлено наличием миоглобина и большого числа митохондрий. Восстановление функции после утомления происходит быстро. Нейромоторные единицы этих мышц состоят из большого числа мышечных волокон.
Быстрые физические волокна окислительного типа. Мышцы, которые преимущественно состоят из волокон этого типа, выполняют быстрые сокращения без заметного утомления, что объясняется большим количеством митохондрий в этих волокнах и способностью образовывать АТФ путем окислительного фосфорилирования. Как правило, число волокон, входящих в состав нейромоторной единицы, в этих мышцах меньше, чем в предыдущей группе. Основное назначение мышечных волокон данного типа заключается в выполнении быстрых энергичных движений.
Быстрые физические волокна с гликолитическим типом окисления. Волокна данного типа характеризуются тем, что АТФ в них образуется за счет гликолиза. Волокна этой группы содержат митохондрий меньше, чем волокна предыдущей группы. Мышцы содержащие эти волокна, развивают быстрое и сильное сокращение, но сравнительно быстро утомляются. Миоглобин в данной группе мышечных волокон отсутствует, вследствие чего мышцы, состоящие из волокон этого типа, называют белыми.
Для мышечных волокон перечисленных групп характерно наличие одной, в крайнем случае нескольких, концевых пластинок, образованным одним двигательным аксоном.
Тонические волокна. В отличие от предыдущих мышечных волокон в тонических волокнах двигательный аксон образует множество синоптических контактов с мембранной мышечного волокна. Развитие сокращения происходит медленно, что обусловлено низкой активностью миозиновой АТФазы. Так же медленно происходит и расслабление. Мышечные волокна данного типа эффективно работают в изометрическом режиме. Эти мышечные волокна не генерируют потенциал действия и не подчиняются закону «все или ничего». Одиночный пресинаптический импульс вызывает незначительное сокращение. Серия импульсов вызовет суммацию постсинаптического потенциала и плавно возрастающую деполяризацию мышечного волокна (наружные мышцы глаз).
- Механизм
сокращения скелетной
мышцы.
Двигательная единица-это мотонейрон с иннервируемым и им мышечными волокнами. Аксон мотонейрона из спинного мозга проходит в составе периферических нервов до мышцы, внутри которой разветвляется на множество концевых веточек. Каждая концевая веточка заканчивается на одном мышечном волокне, образуя нервно –мышечный синапс. Двигательная еденица функционирует как единое морфофункциональное образование.
Скелетная мышца состоит из пучков вытянутых в длину клеток-мышечных волокон, обладающих тремя свойствами: возбудимостью, проводимостью и сократимостью. Отличительной чертой мышечных клеток от клеток, не обладающих свойством сократимости, является наличие саркоплазматического ретикулума. Он представляет собой замкнутую систему внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу. Две термальные цистерны ретикулума вместе с трубочкой так называемую триаду-анатомическую структуру, в зоне которой нервные импульсы, распространяющиеся по поперечным трубочкам вглубь мышечного волокна, запускают процесс выхода ионов кальция из саркоплазматического ретикулума, и, следовательно, всю последующую цепочку изменений, приводящую к развитию сокращения мышцы.
В каждом мышечном волокне содержится до 1000 и более сократительных элементов миофибрилл. Каждая миофибрилла состоит из множества параллельно лежащих толстых и тонких нитей-миофиламентов. Толстые нити состоят из молекул белка миозина, а тонкие из белка актина. В состав тонких нитей, кроме того, входит еще два белка-тропонин и тропомиозин, необходимые для развития процессов сокращения и расслабления мышцы.
В покоящихся мышечных волокнах при отсутствии импульсации мотонейрона поперечные миозиновые мостики не прикреплены к актиновым миофиламентам. Тропомиозин расположен таким образом, что блакирует участки актина, способные взаимодействовать с поперечными мостиками миозина. Тропонин тормозит миозин-АТФ-фазную активность и поэтому атф не расщепляется. Физиологическим контролером напряжения мышц являются ионы кальция. В пассивном состоянии ионы кальция скапливаются в особом депо, из которого выбрасываются под действием нервного сигнала. Впоследствии актин реагирует с миозином, что и обеспечивает напряжение мышцы. Транспортировка ионов кальция осуществляется благодаря энергии АТФ. Количества аденозинтрифосфата, находящегося в мышце, достаточно для поддержания функционирования сократительного механизма лишь в течение нескольких миллисекунд. Поэтому при более продолжительных мышечных напряжениях запускаются иные механизмы. В мышце энергия скапливается в виде креатинфосфорной кислоты. Креатинфосфорная кислота обладает более мощным потенциалом транспорта энергетических фосфатных групп, чем аденозинтрифосфат. Фосфагены в виде креатинфосфорной кислоты восстанавливают аденозинтрифосфат, обеспечивая таким образом доставку энергии, требуемой для мышечного напряжения. Однако в сокращающейся мышце количество креатинфосфорной кислоты быстро тратится, а это истощает и запасы аденозинтрифосфата.
- Роль
ионов кальция. Расслабление
скелетной мышцы
.
Тропонин и тропомиозин, лежащие вдоль актиновой спирали, препятствуют присоединению миозиновых поперечных мостиков к актину. Тропонин - единственный белок в актиновых и миозиновых филаментах поперечнополосатых мышц позвоночных животных, имеющий высокое химическое сродство к Са2+.
Каждый тропониновый комплекс связывает четыре иона кальция. Тропониновые комплексы расположены вдоль актинового филамента через каждые 40 нм, прикрепляясь одновременно к актиновому филаменту и молекуле тропомиозина. В состоянии покоя положение тропомиозина конформационно препятствует соединению головок миозина с актиновым филаментом. Связывая Са2+, тропонин претерпевает конформационные изменения, в результате чего молекула тропомиозина смещается и освобождает дорогу миозиновым поперечным мостикам для прикрепления к актиновым центрам. Следовательно, присоединение Са2+ к тропонину устраняет постоянно существующее препятствие для взаимодействия поперечных мостиков с актином. Таким образом, кальций в мышечных волокнах играет роль внутриклеточного посредника, связывающего процессы возбуждения и сокращения.
Сказанное выше объясняет роль Са2+ в регуляции актин-миозинового взаимодействия в скелетных и сердечной мышце позвоночных животных. В большинстве других мышц роль кальция иная.
Расслабление мышцы вызывается обратным переносом ионов Са2+ посредством кальциевого насоса в каналы саркоплазматического ретикулума. По мере удаления Са2+ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы.
Контрактурой называют стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя. Кратковременная контрактура может развиваться после тетанического сокращения в результате накопления в саркоплазме большого количества Са2+ ; длительная (иногда необратимая) контрактура может возникать в результате отравления ядами, нарушений метаболизма.
- Значение
АТФ.
- Заключение.
Степень укорочения мышцы при сокращении зависит от силы раздражителя, морфологических свойств и физиологического состояния. Длинные мышцы сокращаются на большую величину, чем короткие.Незначительное растяжение мышцы, когда напрягаются упругие компоненты, является дополнительным раздражителем, увеличивает сокращение мышцы, а при сильном растяжении сила сокращения мышцы уменьшается.Напряжение, которое могут развивать миофибриллы, определяется числом поперечных мостиков миозиновых нитей, взаимодействующих с нитями актина, так как мостики служат местом взаимодействия и развития усилия между двумя типами нитей. В состоянии покоя довольно значительная часть поперечных мостиков взаимодействует с актиновыми нитями. При сильном растяжении мышцы актиновые и миозиновые нити почти перестают перекрываться и между ними образуются незначительные поперечные связи.
При тренировке увеличивается объем мышц в результате роста и утолщения мышечных волокон возрастает мышечная выносливость. В мышце повышается содержание гликогена, АТФ и креатинфосфата, ускоряется течение процессов распада и восстановления веществ, участвующих в обмене. В результате тренировки коэффициент использования кислорода при работе мышц повышается, усиливаются восстановительные процессы вследствие активизации всех ферментативных систем, уменьшается расход энергии. При тренировке совершенствуется регуляторная функция центральной нервной системы, и в первую очередь, коры больших полушарий.
и т.д.................
Физиология мышц классификация мышечных волокон
Режимы сокращения мышц
Виды сокращений
Строение мышцы
Физиология, мышц классификация мышечных волокон
Мышечные волокна делят на 3 вида: скелетные, сердечные и гладкие.
Скелетные волокна подразделяются на фазные (они генерируют ПД) и тонические (не способны генерировать полноценный потенциал действия распространяющегося типа). Фазные волокна делятся на быстрые волокна (белые, гликолитические) и медленные волокна (красные, окислительные волокна).
Гладкие мышцы делятся на тонические и фазнотонические. Тонические волокна не способны развивать "быстрые" сокращения. В свою очередь фазнотонические мышцы можно условно разделить на обладающие автоматией - способные к спонтанной генерации фазных сокращений, и на мышцы, не обладающие свойством автоматии.
Основным морфо - функциональным элементом нервно-мышечного аппарата является двигательная единица (ДЕ). ДЕ - это мотонейрон с иннервируемыми им мышечными волокнами. Аксон мотонейрона из спинного мозга проходит в составе периферических нервов до мышцы, внутри которой разветвляется на множество концевых веточек. Каждая концевая веточка заканчивается на одном мышечном волокне, образуя нервно-мышечный синапс. Импульсы, идущие по аксону мотонейрона, активируют все иннервируемые им мышечные волокна. Поэтому ДЕ функционирует как единое морфофункциональное образование.
Функции скелетных и гладких мышц
Скелетные мышцы составляют 40% от массы тела и выполняют ряд важных функций:
1 - передвижение тела в пространстве, 2 - перемещение частей тела относительно друг друга, 3 - поддержание позы, 4 - передвижение крови и лимфы, 5 - выработка тепла, 6 - участие в акте вдоха и выдоха, 7 - двигательная активность как важнейший антиэнтропийный и антистрессовый фактор (тезисы "движение - это жизнь" или "кто много двигается, тот много живет" - имеют реальную материальную основу), 8 - депонирование воды и солен, 9 - защита внутренних органов (например, органов брюшной полости).
Гладкие мышцы обеспечивают функцию полых органов, стенки которых они образуют. В частности, благодаря гладким мышцам осуществляется изгнание содержимого из мочевого пузыря, кишки, желудка, желчного пузыря, матки. Гладкие мышцы обеспечивают сфинктерную функцию - создают условия для хранения содержимого полого органа в этом органе, например, мочу в мочевом пузыре, плод в матке. Важнейшую роль выполняют гладкие мышцы в системе кровообращения и лимфообращения - изменяя просвет сосудов, гладкие мышцы тем самым адаптируют регионарный кровоток к местным потребностям в кислороде, питательных веществах. Гладкие мышцы могут существенно влиять на функцию связочного аппарата, т.к содержатся во многих связках и при своем сокращении меняют состояние данной связочной структуры. Например, ГМК (гладкомышечные клетки) содержатся в широкой связке матки.
Режимы сокращения мышц
Для скелетной мышцы характерны два основных режима сокращения - изометрический и изотонический. Изометрический режим проявляется в том, что в мышце во время ее активности нарастает напряжение (генерируется сила), но из-за того, что оба конца мышцы фиксированы (например, мышца пытается поднять большой груз) - она не укорачивается. Изотонический режим проявляется в том, что мышца первоначально развивает напряжение (силу), способную поднять данный груз, а потом мышца укорачивается - меняет свою длину, сохраняя напряжение, равное весу поднимаемого груза. Так как изотоническое сокращение не является "чисто" изотоническим (элементы изометрического сокращения имеют место в самом начале сокращения мышцы), а изометрическое сокращение тоже не является "чисто" изотоническим (элементы смещения все-таки есть, несомненно), то предложено употреблять термин "ауксотоническое сокращение" - смешанное по характеру.
Понятия "изотонический", "изометрический" важны для анализа сократительной активности изолированных мышц и для понимания биомеханики сердца.
Режимы сокращения гладких мышц. Целесообразно выделить изометрический и изотонический режимы (и, как промежуточный - ауксотонический). Например, когда мышечная стенка полого органа начинает сокращаться, а орган содержит жидкость, выход для которой перекрыт сфинктером, то возникает ситуация изометрического режима: давление внутри полого органа растет, а размеры ГМК не меняются (жидкость не сжимается). Если это давление станет высоким и приведет к открытию сфинктера, то ГМК переходит в изотонический режим функционирования - происходит изгнание жидкости, т.е. размеры ГМК уменьшаются, а напряжение или сила - сохраняется постоянной и достаточной для изгнания жидкости.
Виды сокращений
У скелетной мышцы выделяют одиночное сокращение и суммированное сокращение, или тетанус. Одиночное сокращение - это сокращение, которое возникает на одиночный стимул, достаточный для вызова возбуждения мышцы. После короткого скрытого периода (латентный период) начинается процесс сокращения. При регистрации сократительной активности в изометрических условиях (два конца неподвижно закреплены) в первую фазу происходит нарастание напряжения (силы), а во вторую - ее падение до исходной величины. Соответственно эти фазы называют фазой напряжения и фазой расслабления. При регистрации сократительной активности в изотоническом режиме (например, в условиях обычной миографической записи) эти фазы будут называться соответственно фазой укорочения и фазой удлинения. В среднем сократительный цикл длится около 200 мс (мышцы лягушки) или 30-80 мс (у теплокровных). Если на мышцу действует серия прямых раздражении (минуя нерв) или непрямых раздражении (через нерв), но с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания 2-й фазы, то мышца будет на каждый из этих раздражителей отвечать одиночным сокращением.
Суммированные сокращения возникают в том случае, если на мышцу наносятся 2 и более раздражения, причем всякое последующее раздражение (после предыдущего) наносится либо во время 2-й фазы (расслабления или удлинения), либо во время 1-й фазы (укорочения или напряжения).
В случае, когда всякое второе раздражение попадает в период фазы расслабления (удлинения), возникает частичная суммация - сокращение еще полностью не закончилось, а уже возникло новое. Если подается много раздражителей с подобным интервалом, то возникает явление зубчатого тетануса. Если раздражители наносятся с меньшим интервалом и каждое последующее раздражение попадает в фазу укорочения, то возникает так называемый гладкий тетанус.
Строение мышцы
Скелетная мышца состоит из пучков вытянутых в длину клеток - мышечных волокон, обладающих тремя свойствами: возбудимостью, проводимостью и сократимостью . Отличительной чертой мышечных клеток от клеток, не обладающих свойством сократимости, является наличие саркоплазматического ретикулума. Он представляет собой замкнутую систему внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу. В мембране саркоплазматического ретикулума находятся две транспортные системы, обеспечивающие освобождение от ретикулума ионов кальция при возбуждении и их возврат из миоплазмы обратно в ретикулум при расслаблении мышцы. В механизме освобождения ионов кальция из ретикулума при возбуждении мышечной клетки важную роль играет система поперечных трубочек (Т-система), представляющих собой выпячивания поверхностной мембраны мышечного волокна.
Мышечные волокна имеют диаметр от 10 до 100 мкм и длину от 5 до 400 мм (в зависимости от длины мышцы). В каждом мышечном волокне содержится до 1000 и более сократительных элементов миофибрилл, толщиной 1-3 мкм. Каждая миофибрилла состоит из множества параллельно лежащих толстых и тонких нитей - миофиламентов. Толстые нити состоят из молекул белка миозина, а тонкие - из белка актина.
Расположение миозиновых и тонких актиновых белковых нитей строга упорядочено (рис.4.1). Пучок лежащих в середине саркомера нитей миозина выглядит в световом микроскопе как темная полоска. Благодаря свойству двойного лучепреломления в поляризованном свете (то есть анизотропии) она называется А-диском. По обе стороны от А-диска находятся участки, которые содержат только тонкие нити актина и поэтому выглядят светлыми. Эти изотропные J-диски тянутся до Z-пластин. Благодаря такому периодическому чередованию светлых и темных полос миофибриллы скелетной мышцы выглядят исчерченными (поперечно - полосатыми). Если мышца расслаблена, то в средней части А-диска различается менее плотная Н-зона, состоящая только из толстых миофиламентов. Н-зона не просматривается во время сокращения мышцы. По середине J-диска проходит темная полоска - это Z линия. Участок миофибриллы между двумя Z линиями называется саркомером.
Схема саркомера мышечного волокна и взаимного расположения толстых миозиновых и тонких актиновых миофиламентов.
Z - линии, разделяющие два соседних саркомера; J- изотропный диск; А - анизотропный диск; Н - участок с уменьшенной анизотропностью
Механизмы сокращения мышечного волокна. В покоящихся мышечных волокнах при отсутствии импульсации мотонейрона поперечные миозиновые мостики не прикреплены к актиновым миофиламентам.
При сокращении мышцы длина А-дисков не меняется, J - диски укорачиваются, а Н-зона А-дисков может исчезать (рис.4.3). Эти данные явились основой для создания теории, объясняющей сокращение мышцы механизмом скольжения (теорией скольжения) тонких актиновых миофиламентов вдоль толстых миозиновых. В результате этого миозиновые миофиламенты втягиваются между окружающими их актиновыми. Это приводит к укорочению каждого саркомера, а значит, и всего мышечного волокна.
Молекулярный механизм сокращения мышечного волокна состоит в том, что возникающий в области концевой пластинки потенциал действия распространяется по системе поперечных трубочек вглубь волокна, вызывает деполяризацию мембран цистерн саркоплазматического ретикулума и освобождение из них ионов кальция. Свободные ионы кальция в межфибриллярном пространстве запускают процесс сокращения. Совокупность процессов, обуславливающих распространение потенциала действия вглубь мышечного волокна, выход ионов кальция их саркоплазматического ретикулума, взаимодействие сократительных белков и укорочение мышечного волокна называют "электромеханическим сопряжением". Временная последовательность между возникновением потенциала действия мышечного волокна, поступлением ионов кальция к миофибриллам и развитием сокращения волокна показана на рисунке.
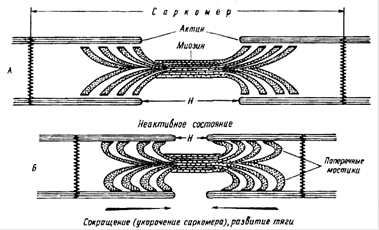
Схема сокращения мышцы
А. Поперечные мостики между актином и миозином разомкнуты. Мышца находится в расслабленном состоянии. Б. Замыкание поперечных мостиков между актином и миозином. Совершение головками мостиков гребковых движений по направлению к центру саркомера. Скольжение актиновых нитей вдоль миозиновых, укорочение саркомера, развитие тяги.
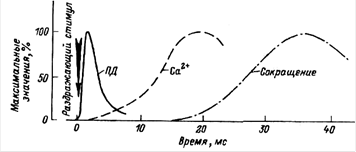
Схема временной последовательности развития возбуждения и сокращения мышцы
При возбуждении волокна Са +2 выходит из цистерн саркоплазматического ретикулума и, следовательно, концентрация его вблизи миофибрилл возрастает. Под влиянием активирующих ионов Са молекула тропонина изменяет свою форму таким образом, что выталкивает тропомиозин в желобок между двумя нитями актина, освобождая тем самым участки для прикрепления миозиновых поперечных мостиков к актину. В результате поперечные мостики прикрепляются к актиновым нитям. Поскольку головки миозина совершают "гребковые" движения в сторону центра саркомера происходит "втягивание" актиновых миофиламентов в промежутки между толстыми миозиновыми нитями и укорочение мышцы.
Источником энергии для сокращения мышечных волокон служит АТФ.
При однократном движении поперечных мостиков вдоль актиновых нитей (гребковых движениях) саркомер укорачивается примерно на 1% его длины. Следовательно, для полного изотонического сокращения мышцы необходимо совершить около 50 таких гребковых движений. Только ритмическое прикрепление и отсоединение головок миозина может втянуть нити актина вдоль миозиновых и совершить требуемое укорочение целой мышцы. Напряжение, развиваемое мышечным волокном, зависит от числа одновременно замкнутых поперечных мостиков. Скорость развития напряжения или укорочения волокна определяется частотой замыкания поперечных мостиков, образуемых в единицу времени, то есть скоростью их прикрепления к актиновым миофиламентам. С увеличением скорости укорочения мышцы число одновременно прикрепленных поперечных мостиков в каждый момент времени уменьшается. Этим и можно объяснить уменьшение силы сокращения мышцы с увеличением скорости ее укорочения.
Поскольку возврат ионов кальция в цистерны саркоплазматического ретикулума идет против диффузионного градиента, то этот процесс требует затрат энергии. Ее источником служит АТФ. Одна молекула АТФ затрачивается на возврат 2-х ионов кальция из межфибриллярного пространства в цистерны. Таким образом, кальций в мышечных волокнах играет роль внутриклеточного посредника, связывающего процессы возбуждения и сокращения.
Регуляция силы сокращения мышц. Для регуляции величины напряжения мышцы центральная нервная система использует три механизма.
1. Регуляция числа активных ДЕ. Чем больше число ДЕ мышцы включается в работу, тем большее напряжение она развивает. При необходимости развития небольших усилий и соответственно малой импульсации со стороны центральных нервных структур, регулирующих произвольные движения, в работу включаются, прежде всего, медленные ДЕ, мотонейроны, которые имеют наименьший порог возбуждения. По мере усиления центральной импульсации к работе подключаются быстрые, устойчивые к утомлению ДЕ, мотонейроны которых имеют более высокий порог возбуждения. И наконец, при необходимости увеличения силы сокращения более 20-25% от максимальной произвольной силы (МП С), активируются быстрые, легко утомляемые мышечные волокна, иннервируемые крупными мотонейронами с самым высоким порогом возбуждения.
Таким образом, первый механизм увеличения силы сокращения состоит в том, что при необходимости повысить величину напряжения мышцы в работу вовлекается большее количество ДЕ. Последовательность включения разных по морфофункциональным признакам ДЕ определяется интенсивностью центральных возбуждающих влияний и порогом возбудимости спинальных двигательных нейронов.
2. Регуляция частоты импульсации мотонейронов. При слабых сокращениях скелетных мышц импульсация мотонейронов составляет 5 - 10 имп/с. Для каждой отдельной ДЕ чем выше (до определенного предела) частота возбуждающих импульсов, тем больше сила сокращения ее мышечных волокон и тем больше ее вклад в развиваемое всей мышцей усилие. С увеличением частоты раздражения мотонейронов все большее количество ДЕ начинает работать в режиме гладкого тетануса, увеличивая тем самым свою силу по сравнению с одиночными сокращениями в 2-3 раза. В реальных условиях мышечной деятельности человека большая часть ДЕ активируется в диапазоне от 0 до 50% МПС. Лишь около 10% ДЕ вовлекаются с дальнейшим возрастанием силы сокращения. Следовательно, при увеличении силы сокращения более 50% от максимальной - основное значение, а в диапазоне сил от 75 до 100% МПС - даже исключительное, принадлежит росту частоты импульсации двигательных нейронов.
3. Синхронизация активности различных ДЕ во времени. При сокращении мышцы всегда активируется множество составляющих ее ДЕ. Суммарный механический эффект при этом зависит от того, как связаны во времени импульсы, посылаемые разными мотонейронами к своим мышечным волокнам. При небольших напряжениях большинство ДЕ работают несинхронно. Совпадение во времени импульсов мотонейронов отдельных ДЕ называется синхронизацией.
Чем большее количество ДЕ работает синхронно, тем большую силу развивает мышца.
Синхронизация активности ДЕ играет важную роль в начале любого сокращения и при необходимости выполнения мощных, быстрых сокращений (прыжки, метания и т.п.). Чем больше совпадают периоды сокращения разных ДЕ, тем с большей скоростью нарастает напряжения всей мышцы и тем большей величины достигает амплитуда ее сокращения.
Функциональные особенности гладких мышц
Гладкие мышцы находятся в стенках внутренних органов и кровеносных сосудов. Регуляция их тонуса и сократительной активности осуществляется эфферентными волокнами симпатической и парасимпатической нервной системы, а также местными гуморальными и физическими воздействиями.
Сократительный аппарат гладких мышц, как и скелетных, состоит из толстых миозиновых и тонких актиновых нитей. Вследствие их нерегулярного распределения клетки гладких мышц не имеют характерной для скелетной и сердечной мышцы поперечной исчерченности. Гладкомышечные клетки имеют веретенообразную форму, длину 50-400 мкм и толщину 2-10 мкм. Они отделены друг от друга узкими щелями (60-150 нм). Возбуждение электротонически распространяется по мышце от клетки к клетке через особые плотные контакты (нексусы) между плазматическими мембранами соседних клеток.
Волокна гладких мышц сокращаются в результате относительного скольжения миозиновых и актиновых нитей, но скорость их сокращения и скорость расщепления АТФ в 100-1000 раз меньше, чем в скелетных мышцах. Поэтому гладкие мышцы хорошо приспособлены к длительному тоническому сокращению без развития утомления.
Гладкие мышцы, обладающие спонтанной активностью, способны сокращаться и при отсутствии прямых возбуждающих нервных и гуморальных воздействий (например, ритмические сокращения гладких мышц кишечника).
Спонтанная активность гладкомышечных клеток связана и с их растяжением, вызывающим деполяризацию мембраны мышечного волокна, возникновение серии распространяющихся потенциалов действия, с последующим сокращением клетки.
Гладкие мышцы, не обладающие спонтанной активностью сокращаются под влиянием импульсов вегетативной нервной системы. Так, в отличие от мышц кишечника, мышечные клетки артерий, семенных протоков и радужки обладают слабой спонтанной активностью, или вообще не проявляют ее. Отдельные нервные импульсы не способны вызвать пороговую деполяризацию таких клеток и их сокращение. Потенциал действия волокна с последующим сокращением возникает лишь при поступлении к нему серии импульсов с частотой 1 имп/с и выше. В гладких мышцах, не обладающих спонтанной активностью возбуждение также передается от одной клетки к последующим через плотные контакты их мембран (нексусы).
Подобно скелетной и сердечной мышцам гладкие мышцы расслабляются, если концентрация ионов кальция снижается ниже 10 -8 моль/л. Однако в связи со слаборазвитым саркоплазматическим ретикулумом и медленным переносом ионов кальция через мембрану клетки, расслабление происходит гораздо медленнее, чем в случае поперечно-полосатых волокон скелетных мышц.







